Биосинтетические пути
Cтраница 1
Биосинтетические пути, очерченные схемой 41, наиболее продуктивно функционируют в наземных растениях. У водорослей и морских беспозвоночных бициклические С20 - изопреноиды лабданового типа синтезируются редко. [1]
Биосинтетические пути кратко описаны в разд. [2]
Биосинтетические пути и соответствующие им катаболические пути контролируются разными регуляторными ферментами. Обычно регуляция соответствующих биосинтетических и катаболи-ческих путей осуществляется координированным образом, реципрокно, так что стимулирование биосинтетического пути сопровождается подавлением катаболи-ческого пути, и наоборот. Более того, биосинтетические пути регулируются обычно на одном из первых этапов. Это избавляет клетку от непроизводительных трат: она не расходует предшественники на синтез тех промежуточных продуктов, которые ей не понадобятся. Мы вновь убеждаемся на этом примере, что принцип экономии лежит в основе молекулярной логики живых клеток. [3]
Биосинтетические пути, ведущие к 20 аминокислотам, встречающимся в белках, и к восьми обычным нуклеотидам нуклеиновых кислот, многочисленны и по большей части довольно сложны. Мы не станем подробно описывать здесь все эти пути; вместо этого мы сосредоточим свое внимание на главных метаболических принципах, лежащих в их основе. Поскольку аминокислоты и нуклеотиды требуются организму в сравнительно небольших количествах, метаболический поток почти никогда не бывает здесь столь большим, как на путях, ведущих в животных тканях к углеводам или жирам. [4]
 |
Заменимые и незаменимые аминокислоты для человека и белой крысы. [5] |
Биосинтетические пути, ведущие к трем родственным аминокислотам - глутамату, глутамину и пролину ( рис. 22 - 1), несложны и у всех форм жизни, по-видимому, одинаковы. Глутамат образуется из аммиака и а-кетоглутарата ( промежуточного продукта цикла лимонной кислоты) под действием L-глутамат-дегидрогеназы. [6]
Биосинтетические пути кратко описаны в разд. [7]
Биосинтетические пути, ведущие к образованию флавоноидов. [8]
Биосинтетические пути регулируются преимущественно по механизму аллостерического ингибирования первого фермента и репрессии синтеза ферментов этого пути конечным продуктом. Регулирование разветвленных биосинтетических путей осуществляется с помощью усложненных вариантов этих же механизмов. [9]
Уникальные биосинтетические пути наблюдаются не только в области ароматических веществ. [10]
Все биосинтетические пути находятся под контролем механизма репрессии конечным продуктом. Точно так же образование большинства анаболических ферментов регулируется путем репрессии их синтеза. Репрессия осуществляется особыми присутствующими в клетке веществами - репрессорами. [11]
Несмотря на то что биосинтетические пути этих соединений тесно взаимосвязаны, факторы, контролирующие их образование, различаются. В случае синтеза рибофлавина у микроорганизмов хорошо изучен генетический контроль и продемонстрирована регуляция через обратную связь конечным продуктом. [12]
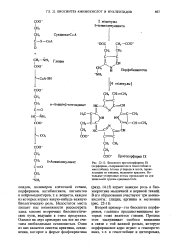
Недостаток места лишает нас возможности рассмотреть здесь многие вторичные биосинтетические пути, ведущие к этим продуктам. Однако на двух примерах мы все же считаем необходимым остановиться. [14]
 |
Различные нуклеозид - и дезокси-нуклеозидтрифосфаты служат теми каналами, через которые энергия АТР направляется на биосинтез тех или иных клеточных компонентов. [15] |
Страницы: 1 2 3