Путь - метаболизм
Cтраница 3
Большое различие в токсичности этой группы смешанных эфи-ров дитиофосфорной кислоты для насекомых и позвоночных объясняется неодинаковыми путями метаболизма препаратов в живых организмах. [31]
Первые две стадии процесса - активация ацетата и образование ацетоацетил - КоА - являются общими для многих путей метаболизма ацетата. [32]
Способность различных органов растений разрушать изосали-пурпозид и флоридзин с образованием различных продуктов связана, по-видимому, с неодинаковыми путями метаболизма этих соединений в хлорофиллоносных и нехлоройиллоносных тканях. [33]
 |
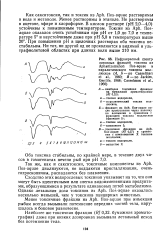
Обмен жирных кислот в печени. [34] |
Как показано на рис. 24 - 11, для жирных кислот, входящих в состав липидов, также характерно несколько путей метаболизма в печени. [35]
Сходство этих водорослевых токсинов указывает на то, что они могут быть идентичными или слегка видоизмененными продуктами, образующимися в результате одинаковых путей метаболизма. [37]
Этот метод может быть с успехом использован не только для оценки остаточных количеств пестицидов в окружающей среде и продуктах питания, но и для изучения путей метаболизма пестицидов в организме, а также в области криминологии и судебной химии. [38]
Однако строгого доказательства этой гипотезы не существовало вплоть до 1950 г., когда сначала Девис [2], а позднее Спринсон [3] и Гибсон [4] и их сотрудники установили путь метаболизма ароматических аминокислот в растениях и микроорганизмах, который теперь известен как шикиматный путь. Природная ( -) - шикимовая кислота ( 7) впервые выделена в 1885 г. Эйкманом из семян плода аниса Illicium religiosum Sieb. Шикимат-ным путем растения и микроорганизмы синтезируют три ароматических аминокислоты: L-фенилаланин ( 10), L-тирозин ( 11) и L-триптофан ( 12); этим же путем растения синтезируют основные ароматические метаболиты. В отличие от растений и микроорганизмов животные не могут использовать этот биохимический путь для синтеза de novo этих аминокислот из углеводных предшественников. [39]
У жвачных основным источником энергии является не глюкоза, а низшие жирные кислоты - уксусная, пропионовая, в меньшей степени масляная и др., образуемые микрофлорой рубца в процессе сбраживания корма. Поскольку путь метаболизма пропионовой кислоты лежит через образование метили ал онил - СоА, то в отсутствие витамина Bi2 она вместе с метилмалоновой кислотой накапливается в крови и вызывает у жвачных потерю аппетита и истощение, столь характерные для дефицита кобальта. Содержание этих кислот в моче жвачных может служить надежным признаком недостаточности кобальта еще задолго до наступления клинических признаков акобальтоза. Таким образом, у жвачных высокая потребность в витамине Bi2 сочетается с низкой эффективностью его синтеза и усвоения, делающими их особенно восприимчивыми к дефициту кобальта по сравнению с другими видами животных. [40]
Значительная часть информации о метаболизме и фарма-кокинетике клоназепама получена в экспериментах с радиоактивным препаратом. Изучение путей метаболизма в организме человека, крыс и собак указывает на сходство протекания процесса в случае клоназепама и нитразепама. Так, у человека и крыс в моче обнаружены практически все перечисленные метаболиты, в то время как у собак только амин и 3-оксипроизводные клоназепама и его восстановленной формы. После внутривенного введения препарата уровень его концентрации понижается через 10 ч, что связано с перераспределением клоназепама в организме людей. Предполагается, что клоназепам и его метаболиты экскретируются с желчью, так как 26 6 - 30 3 % введенной дозы обнаруживается в кале. После перорального приема клоназепама в дозе 1 5 мг через 22 ч с мочой выделяется 12 - 14, после 48 ч - 25 - 29, после 72 ч - 36 - 37 и через 7 дней - около 50 % введенной дозы. В этот период выделение клоназепама почками прекращается. С мочой у взрослых людей выделяется 1 - 2 % неизменного препарата, 40 - 50 % свободных и около 30 % конъюгированных метаболитов. В то же время отмечено присутствие 20 - 30 % свободных, не экстрагируемых метаболитов. Фекалии не содержат исходный препарат, а среди метаболитов обнаружены амин, ацетамид и его 3-оксипроизводное. [41]
Инфекция вызывает изменение путей метаболизма в клетках растения-хозяина в очаге инфекции. Сюда могут включаться и клетки, окружающие очаг инфекции, но самое существенное, что устанавливается локальное взаимодействие между потенциально паразитическим грибом и клетками хозяина, подвергнутыми грибковой инвазии. Изменение путей метаболизма происходит независимо от окончательной реакции ткани хозяина и проявляется в увеличении интенсивности дыхания, синтеза белка и накопления полифенолов, В основном эти изменения непосредственно не связаны с реакциями заболеваний как таковыми, но являются основными результатами инфекции. Фенольные соединения, являющиеся обычными метаболитами незараженных тканей растений, накапливающиеся в очаге инфекции, не обязательно должны играть основную роль в защите растения от инфекции. Известна небольшая группа фенольных соединений, представляющих собой необычные метаболиты, которая не найдена в неинфицированных тканях; вероятно, что эти соединения непосредственно связаны с устойчивостью к заболеванию. Например, пизатин играет основную роль в устойчивости садового гороха к заболеваниям. Медленно идет накопление данных, показывающих, что и у других растений существуют реакции, сходные с приведенными для образования пизатина в горохе. В настоящее время теории Мюллера и Бергера [47], а также Оффорда [46] могут дополнять друг друга; обычно встречающиеся фенольные соединения могут служить предшественниками для токсичных соединений с большей избирательностью, таких, как пизатин, которые образуются только после заражения. [42]
Биология прокариотной клетки как простейшей единицы живого мира рассматривает универсальные, свойственные всем бактериям свойства. Она основывается на знании путей метаболизма в цитозоле; биоэнергетики мембран, механизма синтеза белков на рибосоме, генетики и генома. В отличие от биохимиков и молекулярных биологов, микробиологи имеют дело с микробной клеткой как организованной единой системой, представляющей целостный организм с его реакциями, обусловленными взаимодействием компонентов клетки. [43]
 |
Развитие культуры и синтез вторичного метаболита ( ОП - оптическая плотность суспензии продуцента. [44] |
Вообще, о механизмах переключения путей метаболизма с первичного на вторичный и о физиологической роли вторичных метаболитов в жизни собственного продуцента достоверно известно очень мало. Неизвестно, насколько распространен вторичный метаболизм в природе. Само понятие вторичный метаболит достаточно расплывчатое и многие исследователи его не признают. [45]
Страницы: 1 2 3 4