Распределение - метка
Cтраница 3
Особенностью способа активации на заряженных частицах является то, что ее глубина может регулироваться за счет изменения режима облучения и относительно невелика: общая толщина активируемого слоя обычно не превышает 1 мм, равномерность распределения метки достигается в еще более тонких слоях. [31]
Конечно, прежде чем из распределения метки делать заключения о существовании промежуточной частицы, необходимо убедиться, является ли устойчивым положение метки в исходном веществе и продукте в условиях реакции; только в этом случае распределение метки будет непосредственным результатом рассматриваемой реакции. Это справедливо для вышеприведенного эксперимента, что доказывает существование симметричной промежуточной частицы, хотя она не обязательно должна иметь структуру 16; эта структура была установлена только после дополнительных исследований. [32]
Следует пометить атомы водорода воды ( одновременно за счет обмена будут помечены атомы водорода гидроксильных групп основания, применяемого в качестве катализатора) и в другом опыте - атомы водорода альдегидных групп и проследить за распределением метки ( проще всего - дейтерия) в продуктах реакции. Чтобы выяснить влияние природы катализатора, следует изучить кинетику реакции, вызываемой основаниями различной силы и катионами, различающимися по комплексообразующей способности. [33]
 |
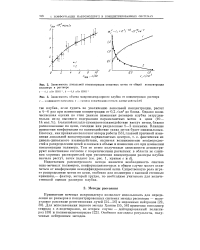
Введение тритиевой метки твердофазным изотопным обменом. [34] |
Протонированные фрагменты или исходно заряженные группы молекул вещества препятствуют изотопному обмену протия на тритий, особенно у соседних углеродных атомов, что приводит к существенному снижению конечной молярной радиоактивности меченого препарата и оказывает значительное влияние на распределение метки. [35]
Аналогично, общее число различных распределений меток в дереве, изображенном на рис. 10.2, должно равняться четырем, так как его центральная вершина может быть помечена четырьмя различными способами, каждый из которых однозначно определяет распределение меток. [36]
Однако существует обширная область обмена веществ, связанная с биосинтезом природных продуктов, изучение которой по сей день сводится к тому, что в питательную среду высших растений и грибов вводят соединения, содержащие радиоактивный углерод, и прослеживают распределение метки. Очень мало известно и о соответствующих ферментативных реакциях; так, например, было выделено всего несколько ферментов, участвующих в биогенезе алкалоидов. [37]
Если можно предположить, что метка, обнаруженная ( после деградации) в положениях 1 4, полностью находится в положении 1, то ввиду совпадения рассчитанных и опытных данных можно считать, что за время жизни карбониево-го иона распределение метки достигает равновесного значения. Однако наблюдается также дальнейшее перераспределение метки в положения 5 и 6, причем перераспределение не достигает равновесия за время жизни карбониевого иона. Реутовым при дезаминировании к-пропиламина ( разд. В данном случае переход метки из положений 2, 3, 1 и 7 в положения 5 и 6 обусловлен переходом водорода из положения 6 в положение 2 или 1, который сопутствует уже рассмотренному переходу углерода в положении 6 из положения 1 в положение 2 и обратно. [38]
Включение 1 - С14 - уксусной и 2 - С14 - мевалоновой кислот в различные каротиноиды показано на срезах плодов и тканей, а включение их в р-каротин - на бесклеточных препаратах из моркови. Распределение метки в каротиноидах, за исключением одного случая, хорошо согласуется с ожидаемым. [39]
В результате реакций, описываемых уравнениями ( 12) - ( 21), происходит превращение пяти молекул глицеральдегид-3 - фосфата в три молекулы рибулозо-5 - фосфата ( фиг. Результаты анализа распределения метки путем расщепления молекул рибу-лозы, меченных после нескольких секунд фотосинтеза в атмосфере с С1402 [8], отражены на фиг. Так был установлен механизм превращения пяти молекул триозофосфата в три молекулы пентозофосфата. [40]
Было показано, что 1 - С14 - ацетат активно включается в аймалин и другие индольные алкалоиды. Данные о распределении метки [43] также согласуются с гипотезой о полиацетатном происхождении нетриптофановой части молекулы указанных алкалоидов. [41]
 |
Зависимость локальной концентрации спиновых меток от общей концентрации полимера в растворе.| Зависимость объема макромолекулярного клубка от концентрации раствора. [42] |
Недостатком рассмотренного метода является необходимость синтеза спин-меченых полимеров, конформация которых в общем случае может отличаться от конформации немодифицированной цепи. Существенную роль играет распределение меток по цепи, особенно для полимеров с высокой степенью прививки - фактор, который трудно, но необходимо учитывать для количественной оценки размеров клубка. [43]
В дальнейшем Блох и его сотрудники продемонстрировали как синтез сквалена из ацетата, так и превращение его в холестерин в печени живой крысы, тем самым сильно подкрепив вывод о том, что именно сквален играет роль промежуточного продукта на пути от ацетата к холестерину. Исходя из характера распределения метки в холестерине разумно предположить, что связующим звеном между холестерином и его предшественником служит дигид-ротритерпен сквален. [44]
Схемы превращения АЛК в гем и хлорофилл, а также схема образования фикобилиновой структуры из порфиринов приведены в гл. Из этих схем следует, что распределение метки в фикобилине будет одинаковым независимо от того, гем или хлорофилл были промежуточными продуктами его биосинтеза. [45]
Страницы: 1 2 3 4