Ключевая реакция
Cтраница 3
Постоянной проблемой при изучении таких реакций является то, что структура, а часто и состав комплекса, участвующего в ключевой реакции, неизвестны. В рассматриваемом случае мы должны допустить, что три карбонильных лиганда расположены таким образом, что комплекс имеет постоянную плоскость симметрии, проходящую через атом железа и центр координированной двойной связи. [31]
Как видно на примере PQQ, в природе может встречаться и ряд других незаменимых пищевых факторов, принимающих участие в ключевых реакциях метаболизма, хотя истинные витаминные свойства их, включая PQQ, пока не раскрыты. [32]
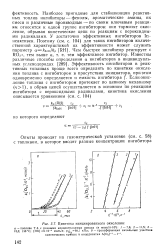 |
Кинетика инициированного окисления. [33] |
Наиболее пригодные для стабилизации реактивных топлив ингибиторы - фенолы, ароматические амины, их смеси и различные производные - по своим ключевым реакциям относятся к одной группе ингибиторов: они тормозят окисление, обрывая кинетические цепи по реакциям с пероксидны-ми радикалами. У достаточно эффективных ингибиторов In-неактивен. [34]
Особого внимания заслуживает наиболее характерная реакция алкиловых эфиров различных производных фосфинистых, фосфонистых и фосфористой кислот-перегруппировка Арбузова103, которая одновременно представляет собой одну из ключевых реакций, необходимых для понимания химии органических соединений фосфора. [35]
Способ гидросилилирования также позволяет напрямую получать анионные, кати-онные и бетаиновые ПАВ из полисилоксановых соединений. Ключевой реакцией является присоединение соответствующего силоксана к промышленно доступному ал-лильному глицидному спирту ( уравн. На практике данное превращение обычно проводят при 40 - 70 С в стеклянном реакторе с использованием около iOppm хлор-платиновой кислоты. Эпоксид полисилоксана получают в виде светло-желтой жидкости, и из-за гидрофобной природы данного материала дальнейшие превращения в ПАВ ( уравн. Возможность химического регулирования молекулярных масс и функциональности полидиметилсилоксано-вых цепей дает огромные возможности химику-синтетику. Четвертичные основания и сульфонаты получают в виде 50 % - ныхводных растворов, а бетаины - с содержанием 30 % - ного активного вещества. [36]
Скорость реакции резко снижается в присутствии окиси углерода, поэтому предполагается, что вначале, как показано, происходит отщепление СО. Ключевой реакцией является раскрытие кольца циклобутена до бутадиена. Ввиду геометрических ограничений, накладываемых на другие циклы, этот процесс должен быть дисротаторным - запрещенным в отсутствие металла. [37]
Уникальный тип циклизации с обоих концов имеет место при биосинтезе соединений с сифолановым скелетом, производные которого встречаются у некоторых губок. Здесь в ключевой реакции принимает участие скваленовый триэпоксид. [38]
Одной из ключевых реакций в этом синтезе является метилирование дезоксиуридинмонофосфата, процесс весьма сложный, протекающий в несколько стадий. Донором дополнительного углеродного атома в С-5-положении является N5, М10 - метилентетрагидрофолиевая кислота. [39]
Этот механизм связан с удалением иона гидрида из изопентана третга-бутильным карбоний-ионом. Такой обмен водород - галоид является ключевой реакцией в механизме, предложенном [68] для объяснения реакции алкилирования. [40]
Как видно из приведенной схемы, синтез инозиновой кислоты начинается с В-рибозо - 5-фосфата, который, как известно, является продуктом пентозофосфатного цикла и на который переносится в необычной реакции пирофосфатная группа АТФ. Таким образом, данная стадия становится ключевой реакцией в синтезе пуринов. На следующей стадии присоединяется вся молекула глицина к свободной NH2 - rpynne 3 - 5-фосфорибозил-амина ( реакция нуждается в доставке энергии АТФ) с образованием глицинамидрибонуклеотида. Затем, на следующей стадии, цепь удлиняется за счет присоединения формильной группы из М5 1ХГ10 - метенил - ТГФК с образованием формилглицинамид-рибонуклеотида. На следующей стадии замыкается пятичленное имидазольное кольцо и образуется 5-аминоимидазолрибонуклеотид, который способен акцептировать СО2 с образованием рибонуклеотида 5-аминоимидазол - 4-карбоновой кислоты. [41]
В табл. 28 приведены эти механизмы, минимальный набор ключевых реакций и выражения для скорости и длины цепи ингибированного окисления. [42]
Собственно химическая стадия процесса, которая представляет собой взаимодейстие субстрата с реагентом в координационной сфере комплекса. Эти стадии, при всем их разнообразии, как правило, включают комбинации ограниченного числа относительно простых стадий, называемых ключевыми реакциями. [43]
Ингибиторами называются вещества, снижающие скорость реакций. Классические исследования, в которых ингибиторные эффекты изучались на изолированных ферментных системах и на метаболических процессах, сыграли исключительно важную роль в установлении строения субстратов, природы тех групп, при помощи которых субстрат связывается с ферментом, а также в выяснении специфичности ферментативных реакций и самого их механизма. В клетке ингибирование ключевых реакций веществами, которые являются продуктами тех же реакций или тех же ( а иногда и других) метаболических процессов, служит тонким регу-ляторным механизмом, обеспечивающим поддержание относительного постоянства состава внутриклеточной среды, а также ответные реакции на изменения во внешней среде. Наконец, избирательное ингибиторное действие ряда природных и синтетических соединений ( антиметаболитов), несомненно, должно быть положено в основу по крайней мере одного из подходов к разработке рациональной фармакологии и химиотерапии. На этом пути открываются две возможности. Вторая возможность заключается в направленном изменении синтеза ферментов. Поскольку сами ферменты являются продуктами метаболизма, их синтезом, очевидно, можно управлять, воздействуя на метаболизм. [44]
После экстракции ДНК подвергают иммобилизации на нитро-целлюлозном или нейлоновом фильтре, расплавлению и гибридизации с известными последовательностями генов, ответственными за синтез тех или иных специфических ферментов. Так, наличие в пробе тотальной ДНК генов, гибридизуемых с геном, кодирующим нитрогеназу, указывает на присутствие в анализируемом сообществе азотфиксаторов, генов метанмонооксигеназы - метанотрофов, генов РуБисКО - автотрофных микроорганизмов, фиксирующих углекислоту через цикл Кальвина. Расшифровано уже несколько десятков генов, кодирующих ключевые реакции тех или иных процессов, и, применяя метод гибридизации, можно делать выводы о наличии определенных микроорганизмов в анализируемой пробе. [45]
Страницы: 1 2 3 4