Регуляция - транскрипция
Cтраница 1
Регуляция транскрипции на уровне последовательности РНК, уже скопированной с генома, происходит за счет белков рестриктаз и лигаз, к-рые разделяют РНК на блоки; нек-рые из них отбрасываются ( интроны), а другие ( экзоны) сшиваются лигазами и поступают в аппарат биосинтеза белков. [1]
Регуляция транскрипции далеко не всегда реализуется по схеме Жакоба и Моно посредством негативного регуляторного фактора-репрессора. В случае фаговых ДНК считывание определенных генов не происходит и в отсутствие каких-либо репрес-соров. Для включения этих генов необходимы позитивные ре-гуляторные факторы. [2]
Регуляция транскрипции далеко не всегда реализуется по схеме Жакоба и Моно посредством репрессора, негативного ре-гуляторного фактора. [3]
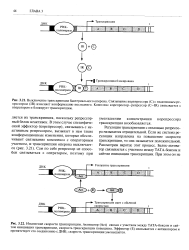
Регуляция транскрипции с помощью репрессо-ра называется отрицательной. Если же система регуляции направлена на повышение скорости транскрипции, то она называется положительной. Рассмотрим вкратце этот процесс. Белок-активатор связывается с участком между ТАТА-боксом и сайтом инициации транскрипции. [5]
Регуляция транскрипции у эукариот отличается от регуляции у прокариот прежде всего тем, что эукариотические РНК-полимеразы при всей сложности их строения сами по себе не в состоянии осуществлять правильную транскрипцию ДНК как в свободном состоянии, так и в составе хроматина, где они реально функционируют. Для инициации транскрипции нужны специальные белковые факторы, разные для трех разных типов эукариотических РНК-полимераз. В настоящее время наиболее детально изучены факторы транскрипции, необходимые для функционирования РНК-полимеразы III, транскрибирующей гены транспортных РНК и рибосомной 5S РНК. [6]
Необходимость регуляции транскрипции в вирусных системах связана с тем, что потребность в разных вирус-специфических белках разная: структурные белки требуются, как правило, в больших количествах, чем белки-ферменты. Кроме того, на ранних стадиях инфекции нужны преимущественно белки, обеспечивающие репликацию генома, а на поздних - белки, принимающие участие в формировании вирионов. Поэтому биологически целесообразно, чтобы разные вирусные гены считывались с разной эффективностью и чтобы эта эффективность изменялась по ходу репродукции вируса. [7]
Механизм регуляции транскрипции заключается в следующем. Белок-репрессор синтезируется в неактивном состоянии в виде прорепрессора. [8]
Сложность регуляции транскрипции в эукариотических клетках проявляется в том, что в них различные типы РНК синтезируются с помощью различных РНК-полимераз. В эукариотическои клетке обнаруживается четыре типа РНК-полимераз, три из которых - РНК-полимеразы I, II и III - локализованы в ядре и одна - в митохондриях. [9]
 |
Схема транскрипции ранних генов фага Т4. [10] |
Система регуляции транскрипции фага Т 4 включает также элементы, которые еще не были описаны. В первую очередь следует Назвать ковалентные и нековалентные модификации РНК-полимера-зы, а также необходимость синтеза вирусной ДНК для включения поздних генов. [11]
Инициация и регуляция транскрипции ДНК у эукариот с участием РНК-полимеразы в большей степени, чем у прокариот, зависит от множества других белков - факторов транскрипции, взаимодействующих с дискретными участками ДНК, образующих сложный эукариотический промотор. В районе промотора, прилегающего к сайту инициации транскрипции ( кэл-сайту), обнаружены участки с характерными нуклеотидными последовательностями ( мотивами), которые оказывают цис-действие на экспрессию близлежащего гена. Эти элементы могут взаимодействовать с РНК-поли-меразой и другими белками-факторами транскрипции. Разные ядерные белковые факторы транскрипции, представляющие собой регу-ляторные белки, способны связываться с теми или иными нуклеотидными последовательностями ДНК, оказывая тем самым влияние на экспрессию разных генов. [12]
Инициация и регуляция транскрипции ДНК у эукариот с участием РНК-полимеразы в большей степени, чем у прокариот, зависит от множества других белков - факторов транскрипции, взаимодействующих с дискретными участками ДНК, образующих сложный эукариотический промотор. В районе промотора, прилегающего к сайту инициации транскрипции ( кзп-сайту), обнаружены участки с характерными нуклеотидными последовательностями ( мотивами), которые оказывают цис-действие на экспрессию близлежащего гена. Эти элементы могут взаимодействовать с РНК-поли-меразой и другими белками-факторами транскрипции. Разные ядерные белковые факторы транскрипции, представляющие собой регу-ляторные белки, способны связываться с теми или иными нуклеотидными последовательностями ДНК, оказывая тем самым влияние на экспрессию разных генов. [13]
Инициация и регуляция транскрипции ДНК у эукариот с участием РНК-полимеразы в большей степени, чем у прокариот, зависит от множества других белков - факторов транскрипции, взаимодействующих с дискретными участками ДНК, образующих сложный эукариотический промотор. В районе промотора, прилегающего к сайту инициации транскрипции ( зл-сайту), обнаружены участки с характерными нуклеотидными последовательностями ( мотивами), которые оказывают цис-действие на экспрессию близлежащего гена. Эти элементы могут взаимодействовать с РНК-поли-меразой и другими белками-факторами транскрипции. Разные ядерные белковые факторы транскрипции, представляющие собой регу-ляторные белки, способны связываться с теми или иными нуклеотидными последовательностями ДНК, оказывая тем самым влияние на экспрессию разных генов. [14]
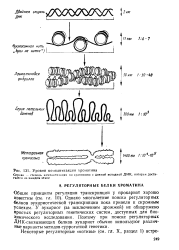 |
Уровни компактизации хроматина. [15] |
Страницы: 1 2 3 4