Репликация
Cтраница 4
Вначале репликация идет по схеме Кэрнса: обнаруживаются 6-молекулы, в которых две уепликационные вилки движутся в противоположных направлениях. Как всегда, движение репликационной вилки, имеющей лидирующую и отстающую цепи, требует сложного ферментативного аппарата. [46]
Вначале репликация вдет по схеме Кэрнса: обнаруживаются 6-молекулы, в которых две репликационные вилки движутся в противоположных направлениях. Как всегда, движение репликационной вилки, имеющей лидирующую и отстающую цепи, требует сложного ферментативного аппарата. [47]
Вначале репликация идет по схеме Кэрнса: обнаруживаются 9-молекулы, в которых две репликационные вилки движутся в противоположных направлениях. Как всегда, движение репликационной вилки, имеющей лидирующую и отстающую цепи, требует сложного ферментативного аппарата. [48]
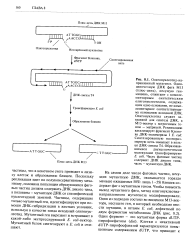
Поскольку репликация идет по полуконсервативному механизму, половина популяции образующихся фаговых частиц должна содержать ДНК дикого типа, а половина - мутантную ДНК со специфической нуклеотидной заменой. Частицы, содержащие только мутантный ген, идентифицируют при помощи ДНК-гибридизации в жестких условиях, используя в качестве зонда исходный Олигонуклеотид. Мутантный белок синтезируют в E. [50]
 |
Пострепликативная репарация. [51] |
Если репликация ДНК произошла до того, как из ДНК были удалены дефекты, может возникнуть показанная на рисунке ситуация, когда одна из цепей ДНК повреждена, а во второй имеется брешь, так как поврежденная цепь не может служить полноценной матрицей. [52]
 |
Пострепликативная репарация. [53] |
Если репликация ДНК произошла до того, как из ДНК были удалены дефекты, может возникнуть показанная иа рисунке ситуация, когда одна из цепей ДНК повреждена, а во второй имеется брешь, так как поврежденная цепь не может служить полноценной матрицей. [54]
 |
Схематическое изображение непрерывного и прерывистого синтеза цепей ДНК при репликации. [55] |
Для репликации ДНК генома человека из одной-единственной точки с подобной скоростью потребовалось бы более 500 ч; вместо этого репликация генома человека происходит в обоих направлениях и одновременно из множества точек ( множество начал репликации), вовлекая от 30000 до 330000 пар оснований. Таким образом, множественность точек начала репликации ДНК, вероятнее всего, является общим правилом для всех клеток эукариот. [56]
Для репликации сведений каталогов в сетевой среде Windows NT Server требуется сервер для репликации ( экспорта) обновленных данных, а также один или несколько компьютеров для получения ( импорта) копий обновленных файлов. [57]
Регуляция репликации основана на том, что PHKI, ингибитор репликации, нестабильна. РНК или, другими словами, количеству копий плазмиды, а концентрация PHKI, которая и определяет скорость взаимодействия с РНКП и частоту инициации, должна быть пропорциональна количеству копий плазмиды и обратно пропорциональна объему клетки. Когда клетки растут и увеличивается их объем, концентрация PHKI падает и становятся возможными новые раунды инициации. После того как произошла репликация, количество копий плазмиды возрастает и одновременно возрастает концентрация PHKI, что подавляет новые инициации до тех пор, пока рост клеток снова не приведет к достаточному разбавлению РНК-ингибитора. [58]
Начало репликации СоШ1 и pBR32 - 2 контролируется небольшими молекулами РНК, которые взаимодействуют с особым участком плазмиды, расположенным поблизости от точки ини - циации репликации. Генетики, стремящиеся максимально увеличить выход белков, кодируемых клонированными в pBR322 генами, получали мутанты с модифицированными РНК. У некоторых таких мутантов число-копий плазмиды составляет более 100, что приводит к существенному увеличению выхода продуктов кодируемых клонированными генами. Интересная особенность репликации GolEl и pBR322 заключается в том, что она полностью обеспечивается ферментами хозяина. [59]
Процессы репликации pBR322 и ColEl, с одной стороны, в хромосомы Е, coli - с другой, различаются тем, что первый нет подавляется хлорамфениколом, который блокирует синтез белка. В присутствии этого соединения репликация хромосом прекращается по завершении уже начавшегося раунда репликации. [60]
Страницы: 1 2 3 4 5