Репликон
Cтраница 1
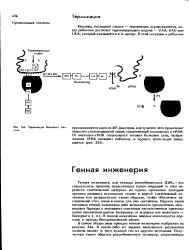
Репликон несет две специфические детерминанты: а) структурный ген ( СГ), контролирующий синтез специфического инициатора, и б) репликатор, или оператор репликации, на который воздействует инициатор, позволяя тем самым копировать ДНК, примыкающую к репликатору ( фиг. [1]
 |
Структура реплицирующейся бактериальной хромосомы. [2] |
Многие репликоны используют, по-видимому, совершенно иную стратегию регуляции собственного синтеза. Этот белок специфически связывается с определенной последовательностью ДНК, многократно повторенной на данном репликоне. Связывание белка-инициатора с одной или несколькими такими последовательностями, находящимися в орид-жине, необходимо для инициации. Одна из последовательностей находится в начале гена белка-инициатора, так что связывание с ней белка подавляет его собственный синтез. Считается, что регуляция репликации осуществляется благодаря сложной конкуренции за белок-инициатор между участком ДНК, необходимым для собственной репрессии, участком ( или участками), необходимым для инициации синтеза ДНК, и другими участками связывания. Хотя подобные репликоны пока еще недостаточно изучены и детальная картина регуляции репликации не ясна, очевидно, что наличие множественных мест связывания ключевого белка инициации репликации позволяет регуляторной системе очень чутко отзываться на изменение копийности репликона. В определенном смысле многократно повторенные участки связывания белка-инициатора, суммарное количество которых пропорционально копийности репликона, аналогичны ранее рассмотренной ингибиторной РНК, концентрация которой также пропорциональна копийности. [3]
Многие мелкие репликоны используют альтернативную стратегию стабильного наследования. Они, по-видимому, не имеют механизма упорядоченной сегрегации, но поддерживаются в высоком числе копий. Высокая копийность обеспечивает относительно стабильное наследование репликона при случайном распределении молекул по дочерним клеткам при делении. Естественно, для этого способа принципиально важным является строгое восстановление ко-пийности, чтобы единственная молекула ДНК успела быстро размножиться до характерного для нее числа копий до начала следующего деления клетки. [4]
Соединение разнородных репликонов ( элементарная генетич. [5]
К наиболее изученным репликонам принадлежат плазмида ColEl и родственные ей плазмиды. Ориджины плазмид этой группы были использованы при конструировании многих векторов для молекулярного клонирования. Репликация ColEl начинается на единственном ориджине и происходит однонаправленно. Для инициации необходимо, чтобы РНК-полимераза синтезировала на ориджине РНК-предшественник затравки, так называемую РНКП. [6]
К наиболее изученным репликонам принадлежат плазмида ColEl и родственные ей плазмиды. Ориджины плазмид этой группы был использованы при конструировании многих векторов для молекулярного клонирования. Репликация ColEl начинается на единственном ориджине и происходит однонаправленно. Для инициации необходимо, чтобы РНК-полимераза синтезировала на ориджине РНК-предшественник затравки, так называемую РНКП. [7]
К наиболее изученным репликонам принадлежат плазмида ColEI и родственные ей плазмиды. Ориджины плазмид этой группы был использованы при конструировании многих векторов для молекулярного клонирования. Репликация ColEI начинается на единственном ориджине и происходит однонаправленно. Для инициации необходимо, чтобы РНК-полимераза синтезировала на ориджине РНК-предшественник затравки, так называемую РНКП. [8]
Следующий уровень образуют репликоны. Каждый репликон представляет собой последовательность нуклеотидов, ограниченную старт - и стоп-символом, или промотором и терминатором, репликации. Как правило, репликоны состоят из нескольких ( до 1000) скриптонов, а также знаков препинания и других функциональных единиц. [9]
Ориджин репликации каждого репликона имеет вполне определенную последовательность ДНК. [10]
 |
Терминация белкового син - присоединяется один из RF-факторов, в результате чего происходит теза гидролиз сложноэфирной связи, соединяющей полипептид и тРНК. [11] |
В какой-либо из заранее выделенных репликонов хозяина вводят in vilro нужный ген из другого источника. [12]
В отличие от оперона репликон начинает функционировать под действием некоторого инициатора, а не индуктора, связывающего репрессор. Инициатор является не негативным, но позитивным регуляторным фактором. Циклическая хромосома ДНК бактерии или фага содержит структурный ген, ответственный за синтез активирующего белка, действующего на ген-репликатор. Возможно, что при этом происходит разрыв кольца и хромосома реплицируется как целое. Многие факты, включая результаты прямых электронно-микроскопических исследований, показывают, что во время деления бактериальной клетки инициатор связан с клеточной мембраной. [13]
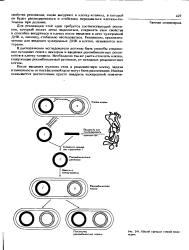 |
Общий принцип генной инженерии. [14] |
Для реализации этой идеи требуется соответствующий репликон, который может легко выделяться, сохранять свои свойства н способен внедряться в клетку после введения в иего чужеродной ДНК и, наконец, стабильно наследоваться. Репликоны, предназначенные для введения чужеродных ДНК в клетки, называются векторами. [15]
Страницы: 1 2 3 4