Репликон
Cтраница 3
Согласно этому механизму, сначала реплицируется только одна из цепей кольцевого репликона, тогда как вторая цепь, оставаясь интактной, вытесняется, образуя петлю. [31]
ДНК, кодирующая ферменты деградации, может быть перенесена из одного репликона в другой. Однако единой единицы транспозиции не существует. [32]
Репликация ДНК происходит только в интерфазе; каждая хромосома имеет по нескольку репликонов; дочерние хромосомы распределяются путем митоза. [33]
Макроэволюционные события [671] приводят к инсерциям, делециям или перераспределению последовательностей ДНК в репликонах. [34]
Инициация репликации ДНК в клетках эукариот регулируется на уровне кластера репликонов, причем число репликонов в кластере изменяется от нескольких единиц до нескольких десятков. [35]
Очень важное свойство мобильных элементов бактерий состоит в том, что они способны вызывать слияние репликонов, или образование коинтегратов. [36]
Мобильные элементы могут не только перемещаться с реп-ликона на репликон, но и вызывать слияние репликонов, образовывать коинтеграт. [37]
Ключевым свойством бактериальных мобильных элементов, обеспечивающим их сохранение, является их способность перемещаться с репликона на репликон. Наличие у бактерий трансмиссивных и мобилизуемых плазмид позволяет транспозонам и IS-элементам не только переходить с плазмиды на плазмиду или из хромосомы на ллазмиду, но и путешествовать из клетки в клетку в составе плазмид. Таким путем мобильные элементы могут распространяться в бактериальных популяциях, даже если не приносят своим хозяевам никаких преимуществ. В этой связи следует упомянуть о явлении иммунности к транспозиции; многие транспозоны и IS-элементы значительно чаще перемещаются на новые репликоны, чем на новое место в составе того репликона, в котором они находятся. Молекулярный механизм этого свойства еще не выяснен, но очевидно, что оно способствует распространению мобильного элемента по максимальному количеству репликонов. [38]
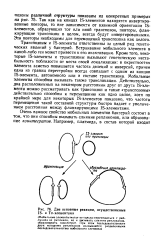 |
Две основные реакции, осуществляемые IS - и Тп-элементами. [39] |
Очень важное свойство мобильных элементов бактерий состоит в том, что они способны вызывать слияние репликонов, или образование коинтегратов. [40]
Мобильные элементы могут не только перемещаться с реп-лнкона на репликон, но и вызывать слияние репликонов. [41]
Ключевым свойством бактериальных мобильных элементов, обеспечивающим их сохранение, является их способность перемещаться с репликона на репликон. Наличие у бактерий трансмиссивных и мобилизуемых плазмид позволяет транспозонам и IS-элементам не только переходить с плазмиды на плазмиду или из хромосомы на ллазмиду, но и путешествовать из клетки в клетку в составе плазмид. Таким путем мобильные элементы могут распространяться в бактериальных популяциях, даже если не приносят своим хозяевам никаких преимуществ. В этой связи следует упомянуть о явлении иммунности к транспозиции; многие транспозоны и IS-элементы значительно чаще перемещаются на новые репликоны, чем на новое место в составе того репликона, в котором они находятся. Молекулярный механизм этого свойства еще не выяснен, но очевидно, что оно способствует распространению мобильного элемента по максимальному количеству репликонов. [42]
Ключевым свойством бактериальных мобильных элементов, обеспечивающим их сохранение, является их способность перемещаться с репликона на репликон. Наличие у бактерий трансмиссивных и мобилизуемых плазмид позволяет транспозонам и IS-элементам не только переходить с плазмиды на плазмиду или из хромосомы на ллазмиду, но и путешествовать из клетки в клетку в составе плазмид. Таким путем мобильные элементы могут распространяться в бактериальных популяциях, даже если не приносят своим хозяевам никаких преимуществ. В этой связи следует упомянуть о явлении иммунности к транспозиции; многие транспозоны и IS-элементы значительно чаще перемещаются на новые репликоны, чем на новое место в составе того репликона, в котором они находятся. Молекулярный механизм этого свойства еще не выяснен, но очевидно, что оно способствует распространению мобильного элемента по максимальному количеству репликонов. [43]
Жакоб и Бреннер [16] считают, что отдельный генетический элемент, например бактериальная хромосома или эписома, образует единицу репликации, или репликон, которая может быть скопирована только целиком. В некоторых отношениях это напоминает оперон при транскрипции ( 284), за исключением того, что, как мы увидим в дальнейшем, он контролируется системой скорее положительной, чем отрицательной регуляции. [44]
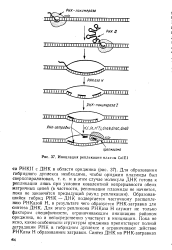 |
Инициация репликации плазмы ColEl. [45] |
Страницы: 1 2 3 4