Роль - белок
Cтраница 2
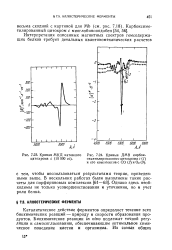 |
Кривая МКД нативного [ IMAGE ] Кривые ДМВ карбок-цитохрома с ( ЮОООгс. метилированного цитохрома с (. /. [16] |
Однако здесь необходимы не только усовершенствования и уточнения, но и учет роли белка. [17]
Нарушение белкового обмена имеет особое значение в нарушении обмена веществ в больном растении, что обусловлено особой ролью белков в жизнедеятельности растений. Белковые вещества участвуют не только во всех метаболических процессах, но и регулируют их на всех уровнях биохимической организации, определяют фенотипические особенности каждого организма и особенности его взаимодействия с паразитом. Поэтому происходящие в больном растении изменения свойств белка влияют, в свою очередь, на ход всех обменных процессов в тканях, а следовательно, и на результаты заражения. [18]
Выше, при обсуждении вопроса о механизме синтеза белков, мы говорили, с одной стороны, о роли белков как шаблонов для этого синтеза, с другой - о роли ферментов. [19]
Белки применяются в медицинской практике, например, в виде лечебной сыворотки. Велика роль белков в легкой промышленности: кожевенная промышленность перерабатывает шкуры животных, текстильная промышленность использует шелк и шерсть, из костей и сухожилий получают клей, желатин. Последний широко используется в пищевой промышленности, в кулинарии. [20]
Понятие о роли белков и полипептидов в живом организме. [21]
Понятие о роли белков и полипептидов в живых организмах. [22]
Медь, первоначально связанная металлотионеином, в дальнейшем включается в церулоплазмин, другие медьсодержащие ферменты и компоненты желчи. Церулоплазмин наряду со своими функциями оксидазы выполняет также роль транспортного белка, переносящего медь на тканевые ферменты, в первую очередь на цитохромоксидазу. В пользу такого допущения свидетельствуют эксперименты, показавшие, что единственным соединением меди, эффективно восстанавливающим активность цитохромоксидазы у крыс, испытывающих недостаточность меди, является именно церулоплазмин. Важную роль в гомеоста-зе меди играют лизосомы гепатоцитов, удаляющие комплекс медь - тионеин из цитоплазмы. Последний превращается в них в водонерастворимую полимерную форму, ошибочно описанную ранее как митохондрокупреин. Лизосомы захватывают частично десиалированный церулоплазмин, который подвергается глубокому протеолизу и затем удаляется из гепатоцитов в желчные протоки. Большая часть асиалоцерулоплазмина ( - 70 %), однако, поступает в желчь, минуя лизосомы. [23]
Мы считаем, что опыты Бренера по переацилированию производных салициловой кислоты можно рассматривать как прототип своеобразной простатической группы белка, создающей такой активный каталитический центр. Работы по переацилированию аминокислот и пептидов с участием ферментов через пептиды оксиаминокислот также подтверждают роль белка как катализатора в реакциях обмена. [24]
 |
Схематическое изображение взаимодействия формилметионил-т РНК и мРНК с 30S субчастицей рибосомы ( а и транслирующей ( функционально активной 70S рибосомой ( б. [25] |
Аналогичные белковые факторы инициации обнаружены также в эука-риотических клетках. Они получены в чистом виде: eIF - 2 состоит из а -, 3 - и у-субъединиц ( мол. Укажем также, что в синтезе белка их роль тождественна роли инициаторных белков у прокариот. Отличительной особенностью синтеза белка у эукариот является, кроме того, наличие среди 10 белковых факторов инициации еще одного белка, названного кэп-связы-вающим. Соединяясь с 5 -участком кэп мРНК, этот белок содействует образованию комплекса между мРНК и 40S рибосомной субчастицей. Необходимо отметить, что до сих пор не раскрыты тонкие молекулярные механизмы участия белковых факторов инициации как у про -, так и у эукариот в сложном процессе синтеза белка. [26]
Более существенные работы исходят из представлений о фазовых переходах в мышце. Сокращение мышцы происходит в результате плавления белковых кристаллов. Активной фазой является релаксация, а не сокращение. Роль сократительного белка состоит в сопротивлении сжатию, роль АТФ - в воздействии на предполагаемый фазовый переход. [27]
Это название не отвечает многочисленным функциям, которые вы полняются белками. Для белков характерно многообразие физиче ских и химических превращений. Бесконечное разнообразие струк туры белков является отражением эволюционного развития живы, существ. Жизненная роль белков связана с повсеместным нахождение. [28]
Качественные особенности белков, как основы жизненных процессов, без сомнения, определяются химическим строением их молекулы. На современном этапе развития учения о белке центральное место в проблеме занимает вопрос о связи структуры индивидуальных белков с их биологическими функциями. Давно отброшены представления о том, что белки - это инертные коллоидные носители низкомолекулярных активных соединений. Ферменты, многие гормоны, токсины, вирусы, антигены, антитела - вот далеко не полный перечень тех важнейших биологически активных тел, свойства которых связаны с их белковой природой. Детальные представления о роли белков в организме начинают вырисовываться только теперь, когда достигнуты принципиальные успехи в деле расшифровки строения индивидуальных белков. Все возрастающее значение идей и методов химии белка как с естественно-научной и философской, так и с чисто практической точки зрения делает необходимой всестороннюю разработку проблемы белка. Одним из предметов такого исследования должна стать история учения о белке. [29]
Устройство мембраны, показанное на рис. 10.2, таково, что белки как бы плавают в липидном море. Их молекулы погружены с двух сторон мембраны на разную глубину в двойной слой подвижных углеводородных хвостов липидов. Имеются белки, проходящие через всю мембрану. Значительная часть поверхности мембраны свободна от белков: так, белки занимают 70 % поверхности мембраны эритроцита и 80 % поверхности мембраны микросомы. Транспорт малых ионов и молекул происходит по каналам в мембранах. В устройстве и функционировании каналов особенно существенна роль белков. [30]
Страницы: 1 2 3