С-конце
Cтраница 2
Все пептиды, у которых на С-конце находится лизин или аргинин, смещаются от диагонального положения, в котором остается только С-концевой пептид исследуемого белка. [16]
 |
Гомологичные аминокислотные последовательности в активных центрах карбоксипептидаз А и В быка. [17] |
С другой стороны, гомологичность последовательностей у С-конца ( 14 остатков) выражена слабее. [18]
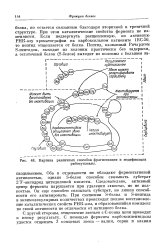 |
Картина различных способов фрагментации и модификации. [19] |
С другой стороны, отщепление звеньев с С-конца цепи приводит к иному результату. [20]
Для полипептидов природного происхождения единственным известным заместителем на С-конце является амидная группа. В функцию амидной группы, по-видимому, входит защита пептидной цепи от атаки карбоксипептидазы или облегчение аккомодации отрицательно заряженного С-конца ( величина р / С а - СО2Н - группы около 3) в неполярном окружении. Как правило, С-концевые модификации значительно более редки, чем N-концевые. [21]
А катализировать последовательное отщепление отдельных аминокислотных остатков от С-конца молекул полипептидов и белков широко используется в научных исследованиях для расшифровки химич. [22]
За пределами 30 - 40 остатков, примыкающих к С-концу, N-концевая часть растущего пептида оказывается свешенной с рибосомы вхокружающую среду. Здесь уже действуют все те факторы, которые определяют спонтанное сворачивание пептида в конформацию, диктуемую условиям среды. Однако необходимо учитывать три обстоятельства, делающих ситуацию отличной от таковой, наблюдаемой при спонтанной ренатурации развернутого белка в опытах in vitro. Во-первых, если рибосома обеспечивала и поддерживала какую-то определенную универсальную конформацию растущего пептида внутри себя, например а-спираль, то сворачивание белка может начинаться не из вытянутого или беспорядочного состояния цепи, а из данной стартовой конформа-ции. Во-вторых, поиск путей сворачивания начинается не с любых и не с разных участков полипептидной цепи, а идет последовательно с N-концевой части цепи. [23]
На первой стадии синтеза аминокислоту, которая будет стоять на С-конце будущего пептида, присоединяют через ее карбоксильную группу к полимеру. При этом нередко используют образование эфирной связи между этой группой и хлорметильной группой полимера. [24]
II), р-нео-эндорфин ( отличается от предыдущего отсутствием на С-конце остатка лизина), динорфин А ( III), динорфин Б ( IV), динорфин-32 ( состоит из остатков динорфина А и динор-фина Б, связанных дипептидом 1 уз - Аг § соотв. С - и Ы - концам), дерморфин ( V), киоторфин ( Н2М - Туг-Аг § - - СООН), казоморфин-5 ( Н2М - Туг-Рго - Рпе-Рго - - О1у - СООН), казоморфии-7 ( отличается от предыдущего наличием на С-конце дополнительно двух аминокислотных остатков Рго-Не и нек-рые др. пептиды. [25]
Полипептид элонгируется ( растет) от М - конца к С-концу. [26]
А ( фА) фЦ, кодируют полилизин с аргинином на С-конце. Это легко понять, если считывание происходит слева направо; в результате такого процесса действительно должен получаться полипептид HaN-Лиз - ( Лиз) га / 3 1 - Аспн-СООИ. [27]
 |
Схема коммуникаций жидкофазного секвенатора белков. [28] |
Принципиально деградация по Эдману при непрерывном повторении циклов возможна вплоть до С-конца. Однако на практике возможно определение последовательности примерно 10 аминокислотных остатков, так как отщепление протекает не всегда количественно, а дополнительное неспецифическое расщепление пептидной цепи вносит ошибку в результаты. [29]
При химической и ферментативной деградации фосфорилированного фермента образуется фосфосерин, связанный по С-концу с остатком гистидина. Аналогичным способом может быть получена и изучена фосфорилированная мутаза фосфоглицериновой кислоты, которая по ряду своих свойств весьма близка к фосфоглюкомутазе. [30]
Страницы: 1 2 3 4