Свертывание - цепь
Cтраница 3
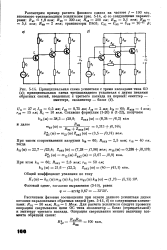 |
Принципиальная схема усилителя с тремя каскадами типа ОЭ. [31] |
Для расчета усилителя следует провести операцию свертывания цепи ОС типа коллектор - база ( отрицательной) к эмиттеру третьего каскада. [32]
Скелетные проволочные модели просты для построения. Если нужно получить приблизительное представление о характере свертывания цепи, можно воспользоваться линейной проволочной моделью с небольшими изгибами в положениях Са. [33]
Исследование специализации белков позволяет устанавливать генеалогию организмов. Как правило, аминокислотная последовательность и характер свертывания цепи в процессе эволюции сохраняются настолько хорошо, что белки даже очень генетически далеких организмов имеют определенное сходство. Поэтому филогенетический анализ, основанный на специализации белков, мэжно рассматривать как один из методов систематики. [34]
Высокий консерватизм укладки цепи связан с ограничениями, налагаемыми динамикой свертывания. Наблюдающаяся у белков тенденция к сохранению способа свертывания цепи в процессе эволюции, по-видимому, не может быть объяснена только на основе сохранения тонкого баланса энергии ( или функции) данного белка. Оба эти условия в значительно большей степени относятся к боковым цепям аминокислот, чем к основной цепи. В связи с этим следует предположить, что укладка цепи сильно коррелирует с динамикой свертывания, которая и налагает основные ограничения на белок. [35]
Скелетные проволочные модели просты для построения. Если нужно получить приблизительное представление о характере свертывания цепи, можно воспользоваться линейной проволочной моделью с небольшими изгибами в положениях Са. [36]
Исследование специализации белков позволяет устанавливать генеалогию организмов. Как правило, аминокислотная последовательность и характер свертывания цепи в процессе эволюции сохраняются настолько хорошо, что белки даже очень генетически далеких организмов имеют определенное сходство. Поэтому филогенетический анализ, основанный на специализации белков, мэжно рассматривать как один из методов систематики. [37]
Высокий консерватизм укладки цепи связан с ограничениями, налагаемыми динамикой свертывания. Наблюдающаяся у белков тенденция к сохранению способа свертывания цепи в процессе эволюции, по-видимому, не может быть объяснена только на основе сохранения тонкого баланса энергии ( или функции) данного белка. Оба эти условия в значительно большей степени относятся к боковым цепям аминокислот, чем к основной цепи. В связи с этим следует предположить, что укладка цепи сильно коррелирует с динамикой свертывания, которая и налагает основные ограничения на белок. [38]
Большинство волокон сверхвысокопрочного / высокомодульного типа являются, как сообщалось, высококристалличными или кристаллизующимися. При изучении малоугловой дифракции рентгеновских лучей не обнаружено свертывания цепи. [39]
Плотность упаковки может служить критерием в предсказании структуры. Плотность упаковки предоставляет возможность независимо проверить результаты расчетного моделирования свертывания цепи ( разд. Этот критерий не сводится только к сравнению со средними плотностями упаковки. [40]
Сравнения топологии слоев подчеркивают важность для структур белков р-складчатых листов. Учитывая трудности перевода средних Са-расстояний в стандартные значимости, следует упомянуть о менее строгом способе сравнения свертывания цепей, основанном на топологии ( 3-структуры и, естественно, применимом только к белкам, содержащим ( 3-складчатые листы. Например, различное расположение листов вдоль цепи или сочленений между параллельными листами на разных сторонах 3-структуры описывает различную топологию 3-структур. [41]
Страницы: 1 2 3