Скорость - инактивация
Cтраница 2
В яде скорпионов содержатся нейротоксины, замедляющие скорость инактивации быстрых натриевых каналов. [17]
В зависимости от термодинамических характеристик процесса наблюдаемая константа скорости инактивации отражает или не отражает равновесия, предшествующего скоростьопределяющей стадии. [18]
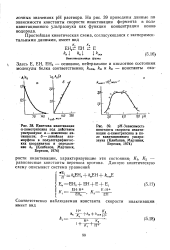
На рис. 39 приведены данные по зависимости константы скорости инактивации фермента в поле кавитационного ультразвука как функции концентрации ионов водорода. [20]
В таблице 12 приведена температурная зависимость эффективной константы скорости инактивации а-трипсина под действием ультразвука. [21]
Помимо уже кратко рассмотренных исследований1660 - 1662 по изучению скорости инактивации бензилпенициллина в водных растворах при различных значениях рН ( см. стр. Для предотвращения такого рода инактивации авторы работы рекомендуют стабилизировать растворы пенициллинов фосфатами, а если это по какой-либо причине невозможно, то применять трубки из другого материала, например поливинилхлорида. В другой работе1ТИ показано, что глицерин способствует разложению пенициллинов водой; это заставляет избегать его присутствия, особенно в больших концентрациях. Появились сообщения 1713 1TU, описывающие уже отмеченный советскими исследователями Г. Е. Казаряном и Э. К. Афри - кяном655 факт стабилизации растворов солей пенициллинов небольшими количествами цитрата натрия. Указано, что последний эффективнее фосфатов. [22]
Принять, что связывание субстрата с ферментом не влияет на скорость инактивации последнего. [23]
Рассмотрим, как с помощью изучения температурной зависимости эффективной константы скорости инактивации можно рассчитать термодинамические параметры АЯ и AS, характеризующие обратимые конформационные изменения активных центров ферментов. [24]
Линейное падение log ( NjN0) со временем показывает, что скорость инактивации пропорциональна числу сохранившихся центров ( как в процессе радиоактивного распада); это указывает, что инактивация осуществляется единым абсорбционным актом и не требует соединенного действия нескольких квант. Арнольд определял абсорбцию ультрафиолетового света у облученной суспензии и подсчитал, что для полной потери активности требуется около 6 квант на каждую молекулу хлорофилла суспензии. Эта цифра ориентировочно указывает, что число восстановительных центров близко к числу хлорофильных молекул. Но он обнаружил, что даже после продолжительного облучения и значительной потери активности, хлорофилл все еще не бледнеет и не изменяется химически. Это напоминает наблюдения над акцептором С02, описанные в главе VIII, где установлено, что хотя этот акцептор не идентичен с хлорофиллом, но он содержится в концентрации, приблизительно равной концентрации зеленого пигмента. [25]
Для этой реакции выведено специальное уравнение и составлена номограмма, позволяющая определять скорость инактивации беизилпенициллина в различных условиях. [26]
С другой стороны, очевидно, что эффекты, приводящие к изменению скорости инактивации, например стабилизации или дестабилизации белка, могут проявляться как на лимитирующей ста-ди реакции, так и на стадии равновесных конформационных изменений, оказывая влияние на константы равновесия. [27]
Кинетика инактивации описывается экспонентой, при этом характеристическое время инактивации определяется константой скорости инактивации этого лабильного фермента. [28]
Была отмечена, таким образом, прямая связь между размерами нативной микробной популяции и скоростью инактивации вирусов. Осталось, однако, неясным, вызвана ли инактивация вирусов токсичными продуктами обмена микроорганизмов или противовирусными энзимами. [29]
 |
Зависимость скорости инактивации v от исходной концентрации субстрата S0 в присутствии защитного вещества Р. [30] |
Страницы: 1 2 3 4