Скорость - рост - популяция
Cтраница 1
Скорость роста популяции зависит от плотности, даже от самой низкой. Можно предположить, что существует порог плотности снизу, при котором особи не взаимодействуют друг с другом. [1]

Скорость роста популяции хищника в этом уравнении зависит от начальной численности ( С) и удельной скорости роста, г - ( К-С) 1К, где К. Относительная скорость в свою очередь зависит от степени недоиспользования среды ( К-С), которую в случае с популяцией хищника можно рассматривать как степень превышения потребностей хищника доступностью жертвы. Однако доступность жертвы и, следовательно, относительная скорость роста популяции хищника часто отражают плотность популяции хищника в некоторый предшествующий период времени ( разд. [3]
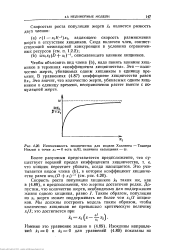 |
Интенсивность хищничества для модели Холдинга - Тэинера Наклон в точке х 0 есть w / D, величина насыщения - w. [4] |
Скорость роста популяции хищников хч такая же, как в (4.89), в предположении, что жертвы достаточно редки. Таким образом, популяция из х жертв может поддерживать не более чем х / ] хищников. [5]
К рассмотрению скорости роста популяции мы вернемся в разд. [6]
Для оценки скорости роста популяции используются методы демографии, а также математического моделирования. [7]
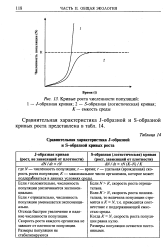 |
Кривые роста численности популяций. [8] |
Когда К N, скорость роста популяции равна нулю. [9]
Аналогично этим задачам рассматривается задача о скорости роста популяции. [10]
При первом типе ( кривая 7) скорость роста популяции уменьшается по мере увеличения плотности. Это широко распространенное явление позволяет понять, почему популяции некоторых животных относительно устойчивы. Во-первых, при увеличении плотности популяции наблюдается снижение рождаемости. Так, в популяции большой синицы при плотности меньше одной пары на 1 га на одно гнездо приходится 14 птенцов; когда же плотность достигает 18 пар на 1 га, выводок составляет менее 8 птенцов. Во-вторых, при увеличении плотности популяции меняется возраст наступления половой зрелости. Например, африканский слон в зависимости от плотности популяции может достигать половой зрелости в возрасте от 12 до 18 лет. Кроме того, этот вид при низкой плотности дает приплод 1 слоненок за 4 года, тогда как при высокой - рождаемость составляет 1 слоненок за 7 лет. [11]
Таким образом, скорость рождаемости, скорость смертности и скорость роста популяции могут быть средними и мгновенными. Далее по тексту подразумевается мгновенная скорость, однако слово мгновенная сознательно опущено, чтобы не перегружать текст. [12]
Именно в постоянном снижении содержания компонентов питательной среды Моно видел причину замедления скорости роста популяции, приводящую к переходу кинетической кривой из экспоненциальной в S-образную. [13]
В целом математическая модель Моно, вызвавшая появление большого числа экспериментальных исследований зависимости скорости роста популяции от концентрации субстрата в питательной среде, а также теоретических представлений в этой области, показала, с одной стороны, важность учета субстрата при анализе роста популяции, а с другой - невозможность сведения закономерностей процесса только к влиянию одного субстрата, да и еще учитываемого в простой схеме ферментативной реакции. Поиски аналогий между кинетикой сравнительно простых ферментативных реакций, проходящих в бесструктурных системах, и зависимостью протекания внутриклеточных биосинтетических процессов от концентрации внеклеточного субстрата хотя и заманчивы, но требуют осторожности. [14]
Основным и главным недостатком разобранных выше моделей является то, что они лишь описывают процесс накопления биомассы или скорость роста популяции в зависимости от условий его осуществления ( куда относят и концентрацию субстрата, и концентрацию биомассы, и продукты метаболизма), а не рассматривают рост как переход субстрата питательной среды в организованную биомассу популяции. Наилучшее совпадение рас-счетных и экспериментальных данных достигается именно в том случае, если рост популяции трактуется не односторонне ( как ограничение со стороны уменьшающегося количества субстрата или как возрастающие в процессе накопления биомассы конкурентные внутривидовые взаимоотношения), а как результат двух противоположно направленных тенденций. С одной стороны, это процесс переработки субстрата питательной среды, результатом чего является увеличение - биомассы живых организмов, регуля-торный аппарат которых, реализуя внутреннюю информацию или информацию микросистем, обеспечивает самовоспроизведение особей. С другой стороны, это тормозящие воздействия процесса, реализующего внешнюю информацию относительно факторов среды обитания, информацию макросистемы. Таким образом, процесс роста популяции не может быть сведен только лишь к учету одного фактора, а является результатом сложных взаимоотношений в популяции. [15]
Страницы: 1 2 3