Скорость - рост - популяция
Cтраница 2
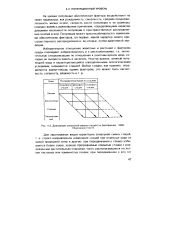 |
Диаграмма зональной смены стаций ( по Бей-Биенко, 1966. [16] |
На уровне популяции абиотические факторы воздействуют на такие параметры, как рождаемость, смертность, средняя продолжительность жизни особи, скорость роста популяции и ее размеры, нередко являясь важнейшими причинами, определяющими характер динамики численности популяции и пространственное распределение особей в ней. Популяция может приспосабливаться к изменениям абиотических факторов, во-первых, меняя характер своего пространственного распределения и, во-вторых, путем адаптивной эволюции. [17]
При отсутствии лимитирующих факторов среды удельная скорость роста равна величине г, которая характеризует свойства самой популяции и называется удельной ( врожденной) скоростью роста популяции или биотическим потенциалом вида. [18]
Вторая концепция - токсическое действие продуктов метаболизма на размножение клеток как фактора взаимодействия особей популяции представлена группой уравнений, предлагающих различную форму связи между скоростью роста популяции и накоплением ингибирующего продукта. Такая неоднозначность математического описания является проявлением относительного характера предполагаемой закономерности роста популяции. Кроме того, практическое использование уравнений, учитывающих ингибирующее действие субстрата, довольно затруднительно в связи со сложностями идентификации токсического компонента среди продуктов метаболизма, а также и возможности количественного учета его концентрации. [19]
Андреварта и Берч [27] также считают, что концепция о роли зависящих от плотности факторов нереалистична, поскольку в ней игнорируется изменение величины г ( показатель скорости роста популяции) во времени, которое может происходить в результате сезонных и других периодических колебаний в среде, а также потому, что эта концепция якобы игнорирует гетерогенность мест, где могут обитать животные. [20]
Описанные модели роста популяции и дифференциальные уравнения предполагают, что все организмы сходны между собой, имеют равную вероятность погибнуть и равную способность к размножению, так что скорость роста популяции в экспоненциальной фазе зависит только от ее численности и не ограничена условиями среды, которые остаются постоянными. Они точно описывают процессы роста и взаимодействия особей в большинстве искусственных и некоторых естественных популяциях. Идеальность всех экологических факторов в исходных условиях предопределила то, что рассматриваемые модели называют идеальными. [21]
Возвращаясь к экспериментальным данным Моно, послужившим основой для создания достаточно наглядной и простой модели роста популяции, следует заметить, что при исследовании влияния увеличивающегося количества субстрата на скорость роста популяции, проводимом на фоне постоянства остальных компонентов питательной среды, не исключена вероятность смены лимитирующего субстрата. То, что это обстоятельство действительно может иметь место, свидетельствуют наблюдения Моно, сделанные им в ходе исследования. [22]
Очевидно, что на самом деле это не так: ближе к центру УСЯОБЯЯ обычно более бяез оприятны вдм в периферии В качестве показателя благоприятности условий можно использовать разньк величины, в ток числг и скорость роста популяции. [23]
Для суспензионного культивирования животных клеток лн-нил ВНК-21 при &09 ч - скорость потребления ими кислорода ( при М; 3 - Ю9 кл / л) не зависит от содержания его в питательной среде и определяется только скоростью роста популяции. [24]
Скорость роста популяции может быть положительной, нулевой и отрицательной. Увеличение ( прибыль) численности происходит в результате рождаемости и имми. [25]
Если N К, скорость роста отрицательна. Если N К, скорость роста популяции равна нулю. [26]
Потребность культуры в кислороде Р, это количество кислорода, которое требуется в единицу времени единице объема культуры для обеспечения растущей популяции необходимым количеством свободной энергии. Потребность культуры в кислороде зависит от скорости роста популяции, которая в свою очередь определяется концентрацией субстрата в питательной среде. [27]
Когда свободная энергия образуется только в процессе дыхания, закономерности размножения микроорганизмов, которые следуют из модели обратимого равновесного автокаталитического роста, могут быть справедливы в том случае, если потребление кислорода соответствует потребности в нем культуры. Если потребление кислорода ниже его потребности, то скорость роста популяции будет ограничиваться скоростью поступления свободной энергии, и ее изменение в этом случае должно подчиняться иным закономерностям, чем те, которые следуют из модели обратимого равновесного автокаталитического роста. [28]
Некоторые исследователи [111, 112] отрицают такую связь. Фенцл [ ИЗ ], признавая кроме положительной зависимости между скоростью роста популяции и запасом питательных веществ в среде существование отрицательной корреляции между скоростью роста и количеством накопившихся микроорганизмов, считает, что до настоящего времени физиологическая сущность этой зависимости еще не ясна: то ли она связана с понижением скорости обмена в околоклеточной микрозоне, то ли вызывается повышением концентрации тормозящих метаболитов, то ли другими причинами. [29]
Вследствие этого уравнение роста популяции не может описать поведение популяции в начальном периоде роста. Установление стационарного состояния характеризуется достижением предельной концентрации промежуточных продуктов, от которых и зависит скорость роста популяции. Именно этот момент рассматривается в приведенных выше уравнениях как точка отсчета процесса роста популяции. [30]
Страницы: 1 2 3