Смерть - клетка
Cтраница 2
Пищевую продукцию можно консервировать путем добавления к ней химических антисептиков, которые в малых концентрациях подавляют развитие микроорганизмов или уничтожают их. Химические консерванты обладают способностью проникать в клетки микроорганизмов, вступать во взаимодействие с веществами протоплазмы, в результате чего наступает смерть клетки. Например, кислоты вызывают гидролиз белков протоплазмы, а спирты их свертывание. Большинство антисептиков ядовито для человека, поэтому их использование строго ограничено. Наиболее распространено применение следующих химических консервантов: этиловый спирт, сернистый ангидрид, бензойнокислый натрий. [16]
В случае летального поражения существует фаза, предшествующая смерти клетки, называемая предлеталь-ной фазой. Если поражающие стимулы, например аноксия, будут ликвидированы в этот период времени, клетка выживет; однако спустя определенный промежуток времени ( критическая точка или момент смерти клетки) устранение поражающего фактора не приводит к восстановлению, в клетке происходят деградация и гидролиз, в конечном итоге достигается физико-химическое равновесие с окружающей средой. Эта фаза называется некрозом. Во время предлеталь-ной фазы происходит ряд принципиальных изменений в зависимости от клетки и типа поражения. [17]
Необходимо сказать, что время должно быть постоянно контролируемым фактором, чтобы оказываемая помощь пострадавшему была действенной. Если же спустя 3 - 4 мин после прекращения дыхания или сердечной деятельности человека ( одновременно или врозь) оказание помощи начато не будет, то, в общем, ее можно уже и не оказывать, так как через 6 - 8 мин уже наступает необратимая биологическая смерть клеток головного мозга и, следовательно, человека. [18]
 |
Поражение клетки. [19] |
Практически вся медицина направлена на предотвращение смерти клетки при таких заболеваниях, как инфаркт миокарда, травма и шок, либо причин, ее вызывающих, как в случае инфекционных болезней и рака. В связи с этим необходимо понимать природу и механизмы этого явления. Смерть клетки классифицировалась как случайная, то есть вызванная токсичными агентами, ишемией и т.п., либо программированной, что происходит во время развития эмбриона, включая образование пальцев и резорбцию хвоста головастика. [20]
 |
Поражение клетки. [21] |
Таким образом, повреждение клетки и смерть клетки играют важную роль как в физиологии, так и патофизиологии. Физиологическая смерть клетки чрезвычайно важна во время эмбриогенеза и развития эмбриона. Исследование смерти клетки во время развития позволило получить важную информацию о протекающих процессах молекулярной генетики, особенно при изучении развития беспозвоночных животных, У этих животных были изучены точное расположение и значение клеток, которым предстоит пережить клеточную смерть, а при помощи классических методов мутагенеза был обнаружен ряд участвующих генов. [22]
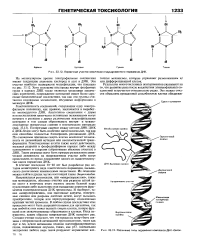 |
Первичные участки химически индуцированного поражения ДНК.| Различные типы поражения комплекса ДНК-белок. [23] |
Генотоксичность соединений, содержащих одну электро-фильную половинку, как правило, заключается в выработке моноаддуктов ДНК. Поперечные сшивки между цепями ДНК-ДНК и ДНК-белок могут быть особенно цитотоксичными, так как они способны полностью блокировать репликацию ДНК. По очевидным причинам смерть клетки исключает возможность ее дальнейшей мутации или неопластической трансформации. Генотоксичные агенты также могут действовать, вызывая разрывы в фосфодиэфирном каркасе либо между основаниями и сахарами ( образовывая абазовые участки) в ДНК. Такие разрывы могут быть прямым результатом химической активности на поврежденном участке либо могут происходить во время ликвидации одного из вышеупомянутых видов поражения ДНК. [24]
Неправильная репликация, или микропоражения, такие как моноаддукты, абазовые участки или разрывы одной цепи могут в конечном итоге вызвать замену базовой пары нуклеотидов либо включение или выпадение коротких фрагментов полинуклеотидов ДНК хромосомы. И наоборот, такие микропоражения, как массивные аддукты, поперечные сшивки или разрывы двойных цепей, могут вызывать приобретение, потерю или перегруппировку относительно крупных частей хромосом. В любом случае последствия этих процессов могут быть разрушительными для организма, так как любой из них может вызвать смерть клетки, потерю функций или злокачественные образования в клетках. Пока неизвестно, каким образом повреждение ДНК вызывает рак. [25]
Разрабатываются новые методы определения клеточного поражения. В более сложных методах используются флуоресцентные зонды для измерения различных межклеточных параметров, таких как выброс кальция, изменение рН и потенциала мембраны. В целом, эти зонды чрезвычайно чувствительны и способны обнаруживать более тонкие изменения клетки, тем самым снижая необходимость использовать смерть клетки в качестве конечной точки. [26]
Drosophila, а также клеток человека и животных, были выявлены предсмертельные гены. Гены некоторых беспозвоночных имеют партнеров у млекопитающих. Например, ген ced - З, необходимый для программирования смерти клетки у С. У Drosophila ген жатки, по-видимому, участвует в создании сигнала, вызывающего запрограммированную смерть клетки. Другие гены, вызывающие смерть, включают Fas в качестве белка мембраны, а также важный ген р53, подавляющий развитие опухолей, который повсеместно консервируется. Ген р53 индуцируется на белковом уровне после поражения ДНК, а после фосфориляции становится фактором транскрипции других генов, таких как gadd45 и waf - 1, участвующих в передаче сигналов о смерти клетки. Другие прямые ранние гены, например c - fos, c - jun, и c - myc, по-видимому, также участвуют в некоторых системах. [27]
Drosophila, а также клеток человека и животных, были выявлены предсмертельные гены. Гены некоторых беспозвоночных имеют партнеров у млекопитающих. Например, ген ced - З, необходимый для программирования смерти клетки у С. У Drosophila ген жатки, по-видимому, участвует в создании сигнала, вызывающего запрограммированную смерть клетки. Другие гены, вызывающие смерть, включают Fas в качестве белка мембраны, а также важный ген р53, подавляющий развитие опухолей, который повсеместно консервируется. Ген р53 индуцируется на белковом уровне после поражения ДНК, а после фосфориляции становится фактором транскрипции других генов, таких как gadd45 и waf - 1, участвующих в передаче сигналов о смерти клетки. Другие прямые ранние гены, например c - fos, c - jun, и c - myc, по-видимому, также участвуют в некоторых системах. [28]
Галогены в живой клетке и организме человека. Химические свойства водорода и галогенов во многом сходны. Казалось бы, галогены, как и водород, могут входить в состав биомолекул и быть активными участниками клеточного метаболизма, но в действительности живая клетка за редчайшими исключениями не содержит галогенсодержащие биомолекулы. Таким образом, химическое сходство галогенов и водорода не настолько велико, чтобы проявляться в химии клетки. Экспериментально показано, что искусственное введение в метаболические циклы живой клетки фтор - и хлорзамещенных аналогов естественных метаболитов блокирует работу этих циклов и приводит к смерти клеток в организме. И тем не менее хлор относится к числу важнейших биогенных элементов, а фтор и иод - к необходимым микроэлементам. [29]
Drosophila, а также клеток человека и животных, были выявлены предсмертельные гены. Гены некоторых беспозвоночных имеют партнеров у млекопитающих. Например, ген ced - З, необходимый для программирования смерти клетки у С. У Drosophila ген жатки, по-видимому, участвует в создании сигнала, вызывающего запрограммированную смерть клетки. Другие гены, вызывающие смерть, включают Fas в качестве белка мембраны, а также важный ген р53, подавляющий развитие опухолей, который повсеместно консервируется. Ген р53 индуцируется на белковом уровне после поражения ДНК, а после фосфориляции становится фактором транскрипции других генов, таких как gadd45 и waf - 1, участвующих в передаче сигналов о смерти клетки. Другие прямые ранние гены, например c - fos, c - jun, и c - myc, по-видимому, также участвуют в некоторых системах. [30]
Страницы: 1 2 3