Нервно-мышечное соединение
Cтраница 1
Нервно-мышечное соединение очень чувствительно к ФОС. Холмс и Робине [49 ] показали, что блокирующие эффекты ТЭПФ, Дфф и зарина ( 10 - 6 М) на различные нервно-мышечные препараты млекопитающих снимаются реактиватором холинэстеразы 2 - ПАМ, и сделали вывод, что блокирующее действие связано с угнетением холинэстеразы. Дуглас и Пантон [17] исследовали влияние ТЭПФ на кошек: малые дозы ( 0 5 мг / кг) повышали чувствительность мышечной концевой пластинки к повторному раздражению; большие дозы ( например, 2 мг / кг) вызывали быструю деполяризацию концевой пластинки. Авторы считают, что эффект был связан исключительно с накоплением ацетилхолина, возникавшим либо местно, либо на более широком пространстве, как следствие угнетения холинэстеразы. Однако наблюдаемые эффекты неожиданно оказались обратимыми - восстановление обычно наступало через 1 час. Хотя оба эти исследования и поддерживают гипотезу о том, что угнетением холинэстеразы можно объяснить описанное действие ФОС, они ни в коей мере не могут считаться безусловно доказательными. [1]
 |
Влияние ингибитора холинэстеразы ( нео-стигмина на длительность постсинаптического потенциала ( потенциала концевой пластинки одиночного мышечного волокна.| Оптимум ( Ор и пессимум ( Ps. [2] |
Если на нервно-мышечное соединение действует какое-либо из этих веществ, постсинаптический потенциал увеличивается по своей амплитуде и резко растягивается по времени. [3]
Нарушение передачи в нервно-мышечном соединении наблюдается в опытах как in vitro, так in situ, при субокщшетальном или системном введении яда кобры В. Л. Юсин, 1950; М. Н. Артемов с соавт. Угнетение нервно-мышечной передачи под действием яда может реализоваться с помощью двух механизмов. Один из них может быть связан с блокирующим действием яда на концевую пластинку, В основе второго может лежать его деполяризующее действие на возбудимые мембраны. Однако при использовании цельного яда трудно отдифференцировать эти два механизма, так как его деполяризующее действие приводит к блокированию распространяющегося возбуждения в нервных проводниках ( Б. Н. Орлов с соавт. [4]
Хорошо известно, что нервно-мышечное соединение позвоночных весьма - чувствительно к ацетилхолину. [5]
Типичное окончание аксона в нервно-мышечном соединении содержит много тысяч таких пузырьков, нз которых под действием одного нервного импульса опоражнивается только несколько сотен. Разработанные в последнее время методики позволяют фиксировать пузырьки в момент слияния нх с пресинаптической мембраной; это достигается быстрым замораживанием ткани в первые миллисекунды после стимуляции нерва ( см. разд. Однако все имеющиеся данные о механизме высвобождения медиатора были первоначально получены в электрофнэнологических экспериментах. [6]
Установлено, что в области нервно-мышечного соединения в больших концентрациях присутствует фермент холинэстераза, способная быстро расщеплять ацетилхолин, выделяющийся в нервном окончании. Значение этого процесса становится ясным, если учесть, что в естественных условиях к мышце поступают быстро следующие друг за другом нервные импульсы и постсинаптическая мембрана, деполяризованная предшествующей порцией ацетилхолина, становится малочувствительной к действию следующей порции. Чтобы идущие друг за другом нервные импульсы могли осуществлять нормальное возбуждающее действие, необходимо к моменту прихода каждого из них убрать предшествующую порцию медиатора. Эту функцию и выполняет холинэстераза. Холин, освобождающийся при расщеплении молекул ацетилхолина, переносится обратно в нервное окончание специальной транспортной системой, существующей в пресинапти-ческой мембране. [7]
Никотиновые рецепторы обнаружены и в нервно-мышечных соединениях. Если произвести аппликацию никотина непосредственно на это соединение, то мышца сократится, иными словами, никотин имитирует эффект ацетилхо-лина. Никотиновые рецепторы имеются в некоторых зонах головного мозга, однако их связь с психотропным действием никотина неясна. Многие из них стимулируют высвобождение дофамина и, следовательно, системы поощрения и удовольствия. Однако поведенческие эффекты никотина по сравнению с другими психотропными средствами очень слабые. [8]
Эффекты кураре наиболее выражены в нервно-мышечных соединениях: мышцы теряют способность сокращаться, и все тело быстро парализуется. Пострадавший умирает от того, что не в состоянии дышать. Впрочем, спасти его может искусственное дыхание, применяемое до тех пор, пока не кончится действие кураре. Парализующие свойства этого препарата используют некоторые южноамериканские индейцы - они смазывают им наконечники стрел, и в результате даже легко раненому животному не удается скрыться от охотника. Курареподобные агенты применяются как миорелаксанты ( расслабите-ли мышц) при проведении операций, поскольку это облегчает работу хирурга. Дыхание в данном случае поддерживается искусственно. [9]
У млекопитающих эти яды действуют на нервно-мышечное соединение, которое чувствительно к ионам и имеет холинергиче-скую природу. [10]
 |
Саркоплазматический ретикулум и Т - система. [11] |
Когда нервный импульс по мотонейрону достигает нервно-мышечного соединения на поверхности мышечного волокна, потенциал действия из области концевой пластинки волной распространяется по Т - системе, передается от нее саркоплазматическому ретикулуму и приводит к высвобождению ионов кальция в саркоплазму. [12]
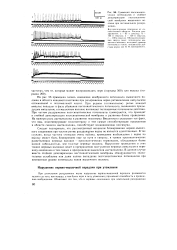 |
Суммация постсинапти-ческих потенциалов и стойкая деполяризация постсинаптиче-ской мембраны мышечного волокна при пессимальном раздражении. [13] |
Следует подчеркнуть, что рассмотренный механизм блокирования нервно-мышечного соединения при частом ритме раздражения нерва не является единственным. В тех случаях, когда частота стимулов очень высока, проведение возбуждения с нерва на мышцу может быть блокировано еще на пути к синапсу, в тонких пресинаптических разветвлениях нервных волокон - пресинаптических терминалях, обладающих более низкой лабильностью, чем толстые нервные волокна. Нарушение проведения в этих тонких нервных волокнах ведет к прекращению поступления нервных импульсов к нервному окончанию и тем самым к прекращению выделения ацетилхолина. В данном случае вместо стойкой деполяризации постсинаптической мембраны обнаруживается значительное ослабление или даже полное выпадение постсинаптических потенциалов при неизменном уровне потенциала покоя мышечного волокна. [14]

Страницы: 1 2 3 4 5