Спектр - белок
Cтраница 1
Спектры белков в области от 1500 до 1800 см 1 можно частично упростить, если изучать их в растворе D. В этих условиях все атомы водорода, присоединенные к азоту, замещаются на дейтерий ( см. раздел Збг), и соответствующие деформационные колебания смещаются в область более низких частот. Полосы поглощения, связанные с деформационными колебаниями D-О - D, также передвинуты в область более низких частот. Таким образом, область от 1500 до 1800 см 1 освобождается от всех полос поглощения за исключением тех, которые обусловлены валентными колебаниями СО-группы. [1]
Спектр ПМР белка состоит из множества перекрывающихся линий. [2]
В спектре ренатурированного белка эти пики занимают более широкую область вплоть до - 0 3т в слабое поле и имеют более сложную структуру. При этом значении рН пики узкие, поскольку протонный обмен с растворителем протекает медленно в шкале времени ЯМР даже для денатурированного белка. Однако этот процесс катализируется основаниями ( см. разд. NH-групп уширяются я исчезают. В условиях частичного обмена вид спектра меняется. Отнесение индивидуальных линий в этой области до сих пор не сделано. [3]
Качественный анализ спектров белков, введенных в пленки ПВС при комнатной и отрицательной температурах, в областях поглощения амид-1 ( 1650 см 1) и амид-2 ( 1540 см 1) позволяет считать, что конформационное состояние пепсина в ПВС соответствует преимущественно а-спиральной форме. [4]
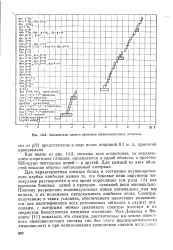 |
Химические сдвиги протонов аминокислотных остатков. [5] |
Для характеристики спектра белка в состоянии неупорядоченного клубка наиболее важно то, что боковые цепи окружены молекулами растворителя и что время корреляции ( см. разд. Поэтому разрешение индивидуальных пиков оптимально, как мы видели, и их положение предсказывать наиболее легко. Спектры, полученные в таких условиях, обеспечивают наилучшие возможности для идентификации всех резонансных сигналов и служат эталонами, с которыми можно сравнивать спектры молекул в их свернутом биологически активном состоянии. [6]
Таким образом, различия в спектрах белков объясняются ионизацией тирозина. Молярный коэффициент поглощения дифференциальной полосы можно получить из спектра поглощения полностью денатурированного щелочью белка по сравнению со спектром нативного белка, приняв во внимание число тирозильных остатков в молекуле или из измерений, выполненных на модельных соединениях. По-видимому, изменения, наблюдаемые при связывании металла с сидерофилинами, действительно, связаны с изменениями в тирозильных остатках, несмотря на то, что не существует зависимости от числа этих остатков в молекуле белка. Тан и Вудворт [64] объяснили свои данные, предположив, что в связывании с железом участвуют два остатка тирозина, а в связывании с медью - один; Ааса и Айзен i [76] сообщили, что при связывании с медью оказываются затронутыми 1 3 остатка тирозина, а Виш-ниа, Вебер и Вернер [75] установили, что в связывании с железом участвуют три таких остатка. Следует указать, что различие в числе спектрофотометрически титруемых остатков тирозина в присутствии металла и без него не обязательно должно рассматриваться как свидетельство их прямой координации к металлу. Возможно также, что наблюдаемые различия вытекают из различий в значениях р / С, изменяющихся в результате конформацион-ных изменений или электростатических эффектов, вызываемых связыванием металла. [7]
 |
Зависимость дифференциальных химических сдвигов а -, Р - и Y ( метальных - протонов валяна от pD ( 100 МГц. [8] |
Сигналы кольцевых протонов гистидинового остатка довольно легко выделить в спектрах белков ( см. гл. [9]
 |
Характеристики полос амид I и амид II. [10] |
Колебательные линии в этих низкочастотных областях действительно удается наблюдать в спектрах КР белков. Соответствующие движения имеют коллективный характер. [11]
Напомним, что аналогичное объяснение было дано выше ( см. § 9.4) при описании спектров ЭПР облученных белков. [12]
Остальные характеристические полосы поглощения высших членов ряда пептидов с увеличением длины цепочки и уменьшением влияния биполярных ионов все больше принимают вид, типичный для спектров белков. [13]
В исследованных пока спектрах строение эмиссионных полос в щелочном растворе почти не зависит от природы белка. Если действительно спектр белка образован наложением спектров тирозина и триптофана, то отсюда следует, что относительное содержание этих компонентов во всех исследованных белках было приблизительно постоянным. [14]
Например, спектры КД белков, свободных и входящих в состав мембранных фрагментов, существенно отличаются: на спектрах белков в составе мембран наблюдаются характерное сглаживание и сдвиг пиков в длинноволновую область. Аналогичные эффекты наблюдаются для спектров ДОВ. Эти эффекты объясняют тем, что пучки с правой и левой круговой поляризации рассеиваются частицами с входящими в их состав оптически активными веществами в неодинаковой степени. Подтверждением такому объяснению служит зависимость формы спектра КД от геометрии расположения детектора ( фотоумножителя) и образца. Приближение образца к детектору в значительной степени устраняет различие в спектрах белков, свободных и входящих в состав мембран. [15]
Страницы: 1 2 3