Спектр - белок
Cтраница 2
Например, спектры КД белков, свободных и входящих в состав мембранных фрагментов, существенно отличаются: на спектрах белков в составе мембран наблюдаются характерное сглаживание и сдвиг пиков в длинноволновую область. Аналогичные эффекты наблюдаются для спектров - ДОВ. Эти эффекты объясняют тем, что пучки с правой и левой круговой поляризации рассеиваются частицами с входящими в их состав оптически активными веществами в неодинаковой - степени. Подтверждением такому объяснению служит зависимость формы спектра КД от геометрии расположения детектора ( фотоумножителя) и образца. Приближение образца к детектору в значительной степени устраняет различие в спектрах белков, свободных и входящих в состав мембран. [16]
Таким образом, сложное строение спектра белков может обусловливаться наложением спектров этих аминокислот, на которые, вероятно, еще налагается спектр фенилаланина, обладающего гораздо более слабым послесвечением со временем жизни менее 0 1 сек. [17]
При денатурации белка в области поглощения 250 - 300 ммк наблюдается небольшой сдвиг ( порядка 1 ммк) в коротковолновую область. Поскольку этот сдвиг весьма мал, спектр белка, подвергшегося денатурации или претерпевшего изменения другого редау обычно сравнивают о спектром натйвного белка, взятого в той же концентрации и растворенного в том же растворителе. [18]
Спектры всех белков имеют большое сходство, но различаются в деталях, что обусловлено, в частности, связыванием малых молекул, свертыванием и развертыванием цепи и другими структурными изменениями. Общее отнесение резонансных сигналов протонов в спектрах белков подобно тому, которое используется для аналогичных малых молекул - аминокислот и пептидов, описанных в гл. Отнесение частот для 20 обычно встречающихся в белках аминокислот приведено на рис. 14.2. Эти данные взяты в основном из тщательно выполненной большой работы Мак-Дональда и Филиппса [11]; сделаны лишь некоторые уточнения, учитывающие отклонения химических сдвигов для аминокислот в длинных полипептидных цепях по сравнению со свободными аминокислотами или короткими пептидами. Следует учитывать, что приведенные значения относятся к белковым цепям в полностью развернутом неупорядоченном состоянии в предположении ( оно почти всегда соблюдается), что отсутствуют взаимодействия между соседними остатками. Это относится к протонам кольца гистидина и метиленовым группам, соседним с амино-группами или карбоксильными группами боковых цепей. Химические сдвиги концевых групп, а также про-стетических групп, таких, как гем-группы, не указаны. Не приводятся также сдвиги протонов групп NH2, ОН, СООН и МН-групп имидазола, поскольку их сигналы обычно сливаются с сигналом от растворителя вследствие быстрого обмена ( см. разд. [19]
Таким образом, в макромолекулах ЯЭО отрицателен и по величине в 2 раза превышает ЯЭО в условиях предельного сужения. Это очень удобно для отнесения сигналов в спектрах белков, особенно если ЯЭО определяется в двумерном варианте ( см. гл. [20]
В совокупности с данными о положении этих сигналов в спектре ЯМР восстановленного белка это может существенно помочь в интерпретации спектра. Дальнейший шаг состоял в определении времен релаксации Tj разрешенных резонансных сигналов в спектре окисленного цитохрома с, что в принципе позволяет оценить расстояния между парамагнитным атомом железа и резонирующими ядрами, как описано выше в этой главе. [21]
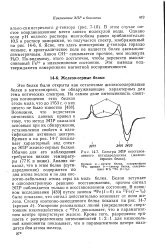 |
Спектры ЭПР восстановленного путидаредоксина ( железо-серного белка. [22] |
Эти белки были открыты как остаточные железосодержащие белки в митохондриях, не обнаруживающие характерных для тема оптических спектров. Кривая а на рис. 14 - 7 показывает характерный вид спектра ЭПР железо-серных белков. Анализ показал, что в этом белке ( пути-даредоксине) содержится по два атома железа и по два кислотно-лабильных атома серы на моль белка. Белок вступает в одноэлектронную реакцию восстановления, причем сигнал ЭПР наблюдается только в восстановленном состоянии. В изящном исследовании [439] путем замены 56Fe ( / 0) на 57Fe ( / 1 / 2) было показано, что сигнал действительно возникает от иона железа. Уширение резонансных линий ( определяемое сверхтонким взаимодействием с 57Fe) доказывает, что в парамагнитном центре присутствует железо. Кроме того, разрешение триплета с распределением интенсивностей 1: 2: 1 для низкополь-ной компоненты показывает, что в парамагнитном центре находятся два атома железа. [23]
Например, спектры КД белков, свободных и входящих в состав мембранных фрагментов, существенно отличаются: на спектрах белков в составе мембран наблюдаются характерное сглаживание и сдвиг пиков в длинноволновую область. Аналогичные эффекты наблюдаются для спектров ДОВ. Эти эффекты объясняют тем, что пучки с правой и левой круговой поляризации рассеиваются частицами с входящими в их состав оптически активными веществами в неодинаковой степени. Подтверждением такому объяснению служит зависимость формы спектра КД от геометрии расположения детектора ( фотоумножителя) и образца. Приближение образца к детектору в значительной степени устраняет различие в спектрах белков, свободных и входящих в состав мембран. [24]
Например, спектры КД белков, свободных и входящих в состав мембранных фрагментов, существенно отличаются: на спектрах белков в составе мембран наблюдаются характерное сглаживание и сдвиг пиков в длинноволновую область. Аналогичные эффекты наблюдаются для спектров - ДОВ. Эти эффекты объясняют тем, что пучки с правой и левой круговой поляризации рассеиваются частицами с входящими в их состав оптически активными веществами в неодинаковой - степени. Подтверждением такому объяснению служит зависимость формы спектра КД от геометрии расположения детектора ( фотоумножителя) и образца. Приближение образца к детектору в значительной степени устраняет различие в спектрах белков, свободных и входящих в состав мембран. [25]
При денатурации белка в области поглощения 250 - 300 ммк наблюдается небольшой сдвиг ( порядка 1 ммк) в коротковолновую область. Поскольку этот сдвиг весьма мал, спектр белка, подвергшегося денатурации или претерпевшего изменения другого редау обычно сравнивают о спектром натйвного белка, взятого в той же концентрации и растворенного в том же растворителе. [26]
Спектр, как и можно было ожидать, очень сильно упростился. Положение сигналов известных протонов, таких, как протон при С-2 в гистидине, не изменилось, что свидетельствует об отсутствии заметных изменений в характере свернутости полипептидной цепи при замещении большинства протонов на дейтерий. В области резонансных сигналов 5 - СН3 - группы метионина около 8 3т можно ясно различить острые резонансные сигналы четырех остатков Мет, которые в спектре недейтерированного белка скрыты большим числом разных пиков в этой области. [27]
Поглощение в этой области почти полностью обусловлено индольными боковыми цепями триптофана и фенольными боковыми группами тирозина. Фенильные боковые группы фенилаланина тоже поглощают излучение в этой области, но их молярное поглощение намного меньше. Относительно малая разница между спектрами белков и спектрами входящих в них боковых цепей, вероятно, означает, что боковые цепи неупорядочены. [28]
Рассмотрим, например, только три спина, расположенные в рад. Такая картина не содержит прямых указаний на то, что А и X являются составными частями одной и тон же спиновой системы, поскольку тот же результат был бы получен для двух двухспиновых систем АМ и М А, если бы М и М имели одинаковый сдвиг. В сложном случае ( особенно в спектрах белков, где эта проблема возникает часто) ш исследования одномерного спектра иногда нельзя определить, имеется лн перекрывание сигналов в некоторой заданной области спектра. [29]
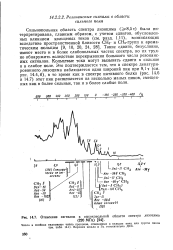 |
Отнесение сигналов в высокопольной области спектра лизоцима. [30] |
Страницы: 1 2 3