Спектр - целлюлоза
Cтраница 2
Такой факт можно рассматривать как указание на то, что тонкая структура v ( ОН) - полосы в спектре целлюлозы обусловлена наличием водородных связей, а не взаимодействием валентных колебаний с другими колебаниями. [16]
Наличие целлюлозы III, по мнению Шпеддинга, характеризуется острым пиком при 3484 см 1 в спектре декристаллизованной целлюлозы, который отсутствует в спектре целлюлозы I. Присутствие целлюлозы III усложняет интерпретацию области гидро-ксилабсорбции. [17]
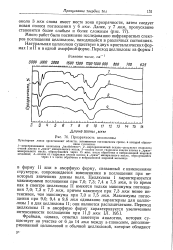 |
Прозрачность целлюлозы. [18] |
Целлюлоза I характеризуется максимумами поглощения при 7 0; 7 3; 7 4 и 7 5 мкм, в то время как в спектре целлюлозы II имеются только максимумы поглощения 7 0; 7 3 и 7 5 мкм, причем максимум при 7 3 мкм менее интенсивен, чем максимумы при 7 0 и 7 5 мкм. Максимумы поглощения при 9 0; 9 4 и 9 6 мкм одинаково характерны для целлюлозы I и для целлюлозы II; они являются расплывчатыми. [19]
Полосы в средней области ИК-спектра целлюлозы II указаны в табл. 6.53. В этой области проявляются в основном те же полосы, что и в спектре целлюлозы I. Различия в интенсивности и положении касаются лишь нескольких полос, связанных с колебаниями ОН - или СН2 - групп. Из этого следует, что обе модификации целлюлозы различаются положением гидроксиметиленовой группы, а также структурой водородных связей. Относительно диффузный характер полос в спектрах целлюлозы II может быть объяснен тем, что эта модификация содержит больше неупорядоченных областей. [20]
Для целлюлозы II, полученной путем регенерации из производных целлюлозы, осаждения растворенной целлюлозы или мерсеризации, область валентных колебаний О - Н - групп резко отличается от соответствующей области спектра целлюлозы I. Это позволяет легко идентифицировать обе структурные модификации. [21]
Основные полосы инфракрасного поглощения для целлюлозы расположены, так же как и для большинства органических молекул, в области 3 - 15 мкм ( 3350 - 667 см 1): В спектре целлюлозы удается разрешить около 30 полос поглощения, обладающих разной интенсивностью. Интенсивность поглощения зависит от изменения дипольного момента в процессе данного колебания. Если молекула состоит из п атомов, то число ее нормальных колебаний составляет Зп - 6; для высокомолекулярных соединений эта величина очень велика. Колебания активны только в том случае, если соответствующие атомы во всех элементарных ячейках смещаются в фазе. Это справедливо только для бесконечной полимерной цепи в кристаллической решетке. Отклонения от идеальной модели приводят к уширению полос спектра и сдвигу частот. Путем рассмотрения одной элементарной ячейки можно предсказать частоты поглощения, а для целлюлозы, основная ячейка которой содержит два остатка целлобиозы ( я 84), можно ожидать 246 нормальных колебаний. Однако взаимодействие между отдельными элементарными звеньями в макромолекуле, по-видимому, во многих случаях ничтожно и число нормальных колебаний должно быть близко к 57 - числу нормальных колебаний для одного элементарного звена. [22]
В спектре хлопка, окисленного периодатом, полоса при 1725 см-1 имеет значительно более низкую интенсивность, чем можно было ожидать. Спектр целлюлозы, в отличие от полиэтилена, имеет только одну широкую полосу в области 1740 см 1 [31, 403, 405], в образование которой вносят вклад как карбоксильные, так и карбонильные группы. Полосы колебания этих групп не удается разделить из-за отсутствия модельных соединений с необходимым окружением групп С О и обычно из-за недостаточно высокого качества образцов. [23]
В работе Марринана и Манна [135] установлено, что дейтерированные модификации целлюлозы I, II, III и IV в области расположения колебаний ОН - - - 3300 см 1 характеризуются различными полосами поглощения. Спектры целлюлозы III и IV меняются в зависимости от того, из какой модификации целлюлозы ( I или II) они приготовлены. [24]
Как видно из рис. 1.10, ИК-спектры двух указанных форм совершенно различны. Спектр целлюлозы II 1ц в области - 3 мкм сохраняет основные особенности спектра целлюлозы II. Это означает, что хотя некоторая перестройка цепей и происходит, по водородные связи, характерные для целлюлоз I и II сохраняются, по крайней мере частично. Однако это не обязательно так, поскольку степень кристалличности образцов мала, и возможно, что часть материала осталась неизмененной. Спектры целлюлоз IVj и IVn очень близки к спектрам целлюлоз I и II соответственно. Из существующего в структурных исследованиях тупика, вероятно, можно выйти путем изучения ориентированных и дейтерированных образцов целлюлоз III и IV. Возможно, будет показано, что при одной и той же упаковке цепей могут существовать водородные связи двух типов. До сих пор такие исследования не были выполнены из-за отсутствия подходящих образцов. [25]
Метод нашел широкое использование в исследовании химических изменений в целлюлозе, ее конфор-мадий ( см. 9.2), водородных связей ( см. 9.3) и др. На рис. 9.1 приведен ИК-спектр целлюлозы, а в табл. 9.1 - интерпретация полос поглощения, т.е. их отнесение к определенным группам атомов, валентные и деформационные колебания которых вызывают возникновение характеристического поглощения. Спектры целлюлоз различного происхождения могут различаться по интенсивности отдельных полос. [26]
Сравнение УФ-спектров отражения целлюлозы и модельных соединений [8 ] показывает, что ацетальная связь не дает существенного вклада в максимум поглощения при 260 нм. Спектры фо-толизованной и фотоокисленной целлюлозы позволяют предположить, что более вероятной причиной появления этого пика служат кетонные карбонильные группы. При фотолизе целлюлозы и амилозы в вакууме содержание кетонных карбонильных групп возрастает, что способствует пожелтению. [27]
Согласно [1594], полосы при 1363, 1264 и 1226 см -, а также смещение от 1110 к 1103 см - - и от 1060 к 1073 см - являются характеристичными для модификации IIIj. Сравнивая спектры целлюлозы II и П1ц, пришли к выводу [662], что они одинаковы, за исключением области поглощения гидроксильных групп. Интенсивность полосы при 900 см - в спектре целлюлозы III является средней между ее значениями для целлюлозы I и II. Все указанные различия очень малы и зависят от степени упорядоченности и методики получения образца. Поэтому трудно проводить анализ кристаллических модификаций III и IV, используя для этого среднюю ИК-область. [28]
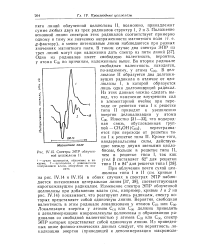 |
Спектры ЭПР облученной целлюлозы II. [29] |
ЭПР наблюдается интенсивная центральная линия [37, 38], соответствующая короткоживущим радикалам. Изменение спектра ЭПР облученной целлюлозы при добавлении влаги ( см., например, кривые / и 2 на рис. IV. Локализация энергии у атомов QD или С ( 4) должна приводить к деполимеризации макромолекулы целлюлозы и образованию радикалов со свободной валентностью у атомов С ( ц или С ( 4), спектр ЭПР которых представляет собой одиночную линию. [30]
Страницы: 1 2 3 4