Терминация
Cтраница 1
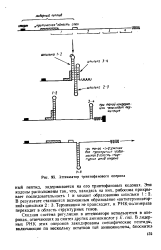 |
Аттенюатор триптофанового оперона. [1] |
Терминации не происходит, и РНК-полимераза переходит в область структурных генов. [2]
Терминация, как уже говорилось в § 5.2, происходит на специальных кодонах-терминаторах UpApA, UpApG и UpGpA. Когда в участок, на котором на рибосоме проходит отбор мономера, попадает такой кодон, связывания какой-либо амино-ацил - тРНК не происходит и синтез полипептидной цепи прекращается. [3]
Терминация представляет собой завершение синтеза полипептидной цепи и освобождение ее от рибосомы. [4]
Терминация в отсутствие факторов, по-видимому, не полностью моделирует этот процесс в том виде, как он протекает в живой клетке. Действительно, на ряде терминаторов очищенная РНК-полимераза завершает синтез молекул РНК, отступя на несколько нуклеотидов от того места, где она завершает его в живой клетке, и с меньшей эффективностью. [5]
Терминации не происходит, и РНК-патнмераза переходит в область структурных генов. [6]
Терминация цепи состоит в том, что по мере движения рибосомы вдоль мРНК возникает сигнал к завершению процесса образования пептида. Такими сигналами служат кодоны УАА, УАГ и УГА. [7]

О терминации полипептида сигнализирует один из трех терминирующих кодонов мРНК, расположенный непосредственно за кодовом последней аминокислоты. Терминирующие триплеты UAA, UAG и UGA не кодируют никакую аминокислоту. Первоначально они были обнаружены при исследовании изменения одного-единственного нуклеотида в некоторых кодонах, соответствующих определенным аминокислотам. [9]
Эффективность терминации зависит от прочности терминатор-ной шпильки в РНК. Это видно из того, что мутации, приводящие к нарушению комплементарного спаривания какого-либо основания в шпильке, ослабляют терминацию, а мутации, восстанавливающие комплементарность, усиливают терминацию. Усиление прочности РНК-ДНК гибрида в районе олигои ослабляет терминацию. [10]
Эффективность терминации зависит от прочности терминагор-ной шпильки в РНК - Это видно из того, что мутации, приводящие к нарушению комплементарного спаривания какого-либо основания в шпильке, ослабляют терминацию, а мутации, восстанавливающие комплементарность, усиливают терминацию. Усиление прочности РНК-ДНК гибрида в районе олигои ослабляет терминацию. [11]
Элонгация и терминация трансляции у про-и эукариот во многом сходны. Первая аминокислота в полипептидной цепи, метионин, отщепляется от тРНК и соединяется с АК2 с помощью пептидной связи. Следующий кодон мРНК спаривается с соответствующим антикодоном тРНК, несущим аминокислоту АКЗ. [12]
Итак, терминация трансляции включает несколько последовательных шагов. [13]
Функционирование факторов терминации, так же как факторов элонгации и одного из факторов инициации, зависит от ГТФ. Для эукарио-тических систем это показано достаточно четко: ГТФ, так же как и его нерасщепляемый аналог, стимулирует кодонзависимое связывание RF с рибосомой, с другой стороны, RF на рибосоме обладает ГТФазной активностью, которая, видимо, нужна для расщепления ГТФ с целью освобождения RF с терминировавшей рибосомы. [14]
В местах р-зависимой терминации РНК-полимераза делает паузы в отсутствие р-фактора, поэтому считается, что роль р-фак-тора заключается в вытеснении РНК из транскрипционного комплекса в местах пауз. Согласно одной из них, фактор движется по синтезируемой РНК, а в местах пауз догоняет РНК-полимеразу и вытесняет РНК-продукт. Другая модель основана на том, что пирофосфат подавляет НТФазную активность р-фактора. Согласно этой модели, р-фактор движется за РНК-полимеразой без отставания, но при нормальной скорости: элонгации ингибируется пирофосфатом, высвобождающимся при синтезе РНК - Активация р-фактора происходит лишь в местах пауз, где синтез цепи РНК временно останавливается, что приводит к прекращению освобождения пирофосфата. [15]
Страницы: 1 2 3 4