Меристематическая ткань
Cтраница 2
В частности, было показано [16], что глубокий покой кончается тогда, когда в меристематических тканях появляются ди - и три-фосфаты нуклеозидов, являющиеся как источниками энергии, так и исходными соединениями для биосинтеза нуклеиновых кислот. Практически полное отсутствие ди - и трифосфатов нуклеозидов в глубоком покое может быть связано, во-первых, с тем, что запас этих соединений был исчерпан растением в результате усиленного морфогенеза в момент формирования клубней на материнском растении. Второй причиной может быть то, что в покоящемся органе блокировано образование макроэргических соединений в процессе окислительного фосфорилирования. [16]
Калий содержат все органы растений в ионном виде, но накапливается он в листьях и меристематических тканях. Не связываемый конституционными веществами, калий перемещается из отмирающих органов к молодым и используется вторично. В цитоплазме калий образует нестойкие соединения с белками, способствует их синтезу, участвует в фотосинтезе. При недостатке его фотосинтез ослабевает, а дыхание растения усиливается. При хорошем калийном питании ускоряется передвижение воды, уменьшается транспирация, повышается засухоустойчивость растений. [17]
Калий содержат все органы растений в ионном виде, но накапливается он в листьях и меристематических тканях. Не связываемый конституционными веществами, калий перемещается из отмирающих органов к молодым и используется вторично. В протоплазме калий образует нестойкие соединения с белками, способствует их синтезу, участвует в фотосинтезе. При недостатке его фотосинтез ослабевает, а дыхание растения усиливается. При хорошем калийном питании ускоряется передвижение воды, уменьшается транспирация, повышается засухоустойчивость растений. [18]
Исследуется изменение содержания химических компонентов в отдельных частях растений, в которых происходит быстрый рост благодаря наличию больших количеств меристематических тканей. [19]
По флоэме гербициды передвигаются в корневую систему, в генеративные органы, накапливаются в зонах активного роста, в меристематических тканях, где и вызывают глубокие нарушения физиологических процессов, приводящих к гибели растений. [20]
Охлаждение суспензионных, каллусных и различных организованных культур проводят обычно медленно ( 1 - 2 С в минуту), меристематические ткани, напротив, охлаждают очень быстро ( от 50 до более чем 1000 С в минуту); также быстро проводят завершающее охлаждение изначально медленно охлаждаемы-х образцов. [21]
В большинстве случаев верхушечные ( длиной 1 - 5 мм) и пазушные побеги заражены растительными вирусами, поэтому прибегают к использованию так называемой культуры верхушечных меристематических тканей, когда отрезают лишь 0 3 - 0 5 мм верхушки, содержащей меристему и 1 - 2 листовых примордия с последующей выдержкой при 30 - 40 С в течение 1 5 -: 3 месяцев. Этот метод удачно использован и используется для оздоровления хозяйственно ценных растений. [22]
К таким активным меристемам относятся клубеньки в период интенсивного роста и формирования бактероидной ткани, поэтому можно предположить, что гербициды - производные феноксиуксусной кислоты передвигаются и аккумулируются в меристематической ткани клубеньков. [23]
Известно, что ауксины и нуклеиновые кислоты имеют одну и ту же сферу влияния: и метаболизм ауксинов, и обмен нуклеиновых кислот и их метаболитов достилают наибольшего напряжения в меристематических тканях точек роста. [24]
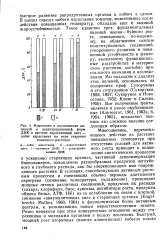 |
Изменения в соотношении на-тивной и денатурированной форм ДНК в системе нарастающий лист - побег израстания по мере старения. [25] |
В условиях, способствующих активным ростовым процессам и сменяющих обычно периоды длительного перегрева ( снижение температуры, увлажнение), происходят мощные оттоки различных соединений, продуктов их распада ( сахара, хлорофилл, биос, ауксины, ингибиторы роста, продукты обмена белка, АТФ, НК) из-за ослабления их связи с клеткой, снижения аккумулятивных сил в стареющих органах ( Engel-brecht, Mothes, 1960, 1964) к физиологически более активным центрам, к возникающим новым меристематическим тканям. Резко меняются при этом конкурентные отношения между стареющими и молодыми органами в пользу последних. [26]
Убедительно демонстрируется с помощью радиоизотопа и далеко не равномерное распределение поступившего фосфора между органами и тканями растения. Активно растущие клетки меристематической ткани корней и листьев получают его в сотни и даже тысячи раз больше, чем те образования, деление клеток которых уже прекратилось. [27]
Этиолированные проростки, зеленеющие на свету, представляют собой удобную синхронную систему, на примере которой можно изучать развитие хлоропластов, однако в ходе нормального образования хлоропластов в тканях растений, растущих на свету, процесс позеленения не происходит. В нормальных условиях в меристематических тканях развиваются небольшие пропластиды, которые в конечном итоге и становятся хлоропластами. Такое превращение происходит без промежуточного образования зтиопластов. Развитие хлоропластов из пропластид в растениях, растущих на свету, изучать довольно трудно, и поэтому пока нет практически никакой информации о синтезе пигментов и его регуляции в подобных системах. У растений разных видов или растущих в условиях разной освещенности отмечены вариации в размерах ССК и в составе входящих в него пигментов. [28]
Более высокую чувствительность двудольных растений к 2 4 - Д можно отчасти объяснить тем, что они имеют большую поверхность листьев, через которые идет поглощение гербицида, и менее защищенное положение точек роста, а значительную устойчивость однодольных, например трав, включая зерновые, тем, что они отличаются небольшой площадью поверхности, плохой смачиваемостью листьев и защищенным положением ростовой точки, находящейся во влагалище листа. Во время интенсивного роста, когда в меристематической ткани происходит интенсивный обмен веществ и ростовая точка обнажается ( у зерновых до достижения ими высоты около 10см и затем вновь во время колошения), однодольные растения также становятся чувствительными к гербицидам типа ростовых веществ. [29]
Результаты, полученные при исследовании последовательных отрезков корня кукурузы, особенно показательны в этом отношении. В первом отрезке ( 2 - 3 мм), который состоит главным образом из меристематической ткани, в обменном фонде в норме процентное содержание каждой кислоты выше, чем в первом односантиметровом отрезке, в котором уже появляется некоторая вакуолизация. В третьем односантиметровом отрезке, в котором наблюдается еще большая вакуолизация, процентное содержание кислот в обменных фондах ниже, чем в первом односантиметровом отрезке. Эти данные находятся в соответствии с представлением, согласно которому по мере развития вакуолизации все большее количество кислот поступает в вакуоли, становясь тем самым менее доступными для действия ферментов цитоплазмы. Эти данные также говорят в пользу представления о том, что в тех растительных клетках, в которых происходит концентрирование кислот, эти последние находятся главным образом в вакуолях, где они оказываются до некоторой степени изолированными от цито-плазматических ферментов. В этой связи чрезвычайно показателен тот факт, что в животных клетках, которым не свойственна вакуолизация, кислоты не накапливаются. [30]
Страницы: 1 2 3 4