Топоизомераза
Cтраница 2
 |
Смена фаз у сальмонеллы. [16] |
И в этом случае Gin-белок действует подобно топоизомеразе, самостоятельно осуществляя и разрыв, и перекрестное лигирование молекул ДНК. В составе G-сегмента имеются гены U и S, кодирующие белки отростка фага, причем в одной ориентации G-сегмента работает один набор этих генов, а в противоположной ориентации - другой ( рис. 72), поскольку G-сегмент транскрибируется только в одном направлении. Salmonella typhimurium и размножаться на этих бактериях, при противоположной ориентации фаг может заражать другие бактерии кишечной группы: Citrobacter, Shigella, Erainia, Serratia. Таким образом, наличие инвертируемого сегмента позволяет фагу Ми существенно расширить круг хозяев. Индивидуальную бактериальную клетку заражает фаговая частица с вполне определенной ориентацией G-сегмента, и при литическом размножении фага эта ориентация преимущественно сохраняется. Но при длительном существовании фага в лизогенных клетках в виде профага происходят множественные акты инверсии, в результате разные лизогенные клетки содержат профагн с различной ориентацией G-сегмента. Рекомбинационный механизм переключения активности генов используют не только фаги. [17]
Аналогичный прием был использован на заключительном этапе очистки топоизомеразы I из зародышей пшеницы. [18]
 |
Схема возможной организации репликативной вилки. [19] |
Нужны также белки, участвующие в инициации репликации, и топоизомеразы. [20]
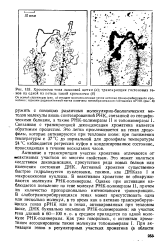 |
Хромосома типа ламповой щетки ( а. транскрипция гистоновых ге - - нов на одной из петель такой хромосомы ( б. [21] |
РНК, связанной со специфическими белками, а также РНК-полимеразы II и топоизомеразы 1 Связанная с транскрипцией деконденсация хроматина является обратимым процессом. Это легко прослеживается на генах дрозофилы, которые активируются при тепловом шоке: при понижении температуры с 37 С до нормальной для дрозофилы температуры 24 С наблюдается регрессия пуфов в конденсированное состояние, происходящая в течение нескольких часов. [22]
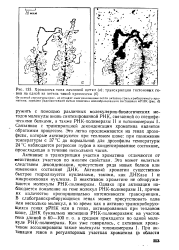 |
Хромосома типа ламповой щетки ( а. транскрипция гистоновых ге - - нов на одной из петель такой хромосомы ( б. [23] |
РНК, связанной со специфическими белками, а также РНК-полимеразы II и топоизомеразы К Связанная с транскрипцией деконденсация хроматина является обратимым процессом. Это легко прослеживается на генах дрозофилы, которые активируются при тепловом шоке: при понижении температуры с 37 С до нормальной для дрозофилы температуры 24 С наблюдается регрессия пуфов в конденсированное состояние, происходящая в течение нескольких часов. [24]
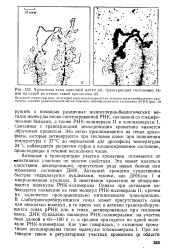 |
Хромосома типа ламповой щетки ( а. транскрипция гистоновых генов на одной из петель такой хромосомы ( б. [25] |
РНК, связанной со специфическими белками, а также РНК-полимеразы II и топоизомеразы 1 Связанная с транскрипцией деконденсация хроматина является обратимым процессом. Это легко прослеживается на генах дрозофилы, которые активируются при тепловом шоке: при понижении температуры с 37 С до нормальной для дрозофилы температуры 24 С наблюдается регрессия пуфов в конденсированное состояние, происходящая в течение нескольких часов. [26]
В последнее время появились данные, свидетельствующие о том, что и работа топоизомераз типа I также идет путем образования разрывов и протаскивания нити через образовавшуюся щель. [27]
Биологическая важность топологических особенностей, которые имеют такого рода структуры, ясна уже из факта существования топоизомераз - присутствующих в живых клетках особых ферментов, способных осуществлять взаимопересечение участков как однонитевой, так и двунитевой ДНК. Среди этих ферментов есть даже такие ( ДНК-гираза), которые ценой затраты энергии способны целенаправленно переводить ДНК в термодинамически невыгодное топологическое состояние, перекручивая нити двойной спирали. [28]
Механизм действия этих препаратов состоит в ингибирова-нии активности бактериальных ферментов - топоизомераз: то-поизомеразы II ( ДНК-гиразы) и топоизомеразы IV. Эти ферменты контролируют структуру и функции ДНК бактерий. Вследствие нарушения процессов репликации хромосомной и плазмидной ДНК бактерии погибают. То есть в результате действия фторхинолонов развиваются бактерицидные эффекты. [29]
Таким образом, в репликации ДНК помимо ДНК-полимеразы принимает участие большой набор ферментов: геликаза, праймаза, РНКаза Н, ДНК-лигаза и топоизомераза II. Этим список ферментов и тем более белков вообще, существенных для матричного биосинтеза ДНК, не исчерпывается. Полная картина еще не выяснена. В качестве еще одного достаточно хорошо изученного белка, необходимого для полноценного функционирования системы ферментов, работающих в вилке репликации, можно привести белок, специфично и кооперативно связывающийся с однонитевыми ДНК. [30]
Страницы: 1 2 3 4