Участие - белок
Cтраница 2
Формы жизни, возникшие на белковой основе, были неустойчивыми из-за отсутствия системы передачи информации, использующей свойства нуклеиновых кислот, а генная жизнь не могла прогрессивно эволюционировать без участия белков, обладающих каталитическими свойствами. Как произошло возникновение формы жизни, в основе которой лежат белки и нуклеиновые кислоты, пока не известно. Ясно только, что встреча обоих типов соединений положила начало пути эволюции, на котором произошло формирование механизмов синтеза белка и нуклеиновых кислот и кодовых взаимодействий между обоими механизмами. [16]
Различная интенсивность распада жиров, углеводов и белков приводит к изменению доли участия их в газообразовании, при этом доля участия жиров, как правило, снижается за счет увеличения доли участия белков, что оказывает прямое влияние на изменение состава газа и иловой жидкости. [17]
Все процессы, протекающие в бактериальной клетке, - образование аминокислот, нуклеоти-дов и других важных метаболитов, репликация, транскрипция, трансляция, катаболизм, высвобождение энергии, реакции на внешние воздействия - требуют участия белков. Однако энергетических ресурсов клетки не хватает для одновременного осуществления транскрипции и трансляции ( экспрессии) всех структурных генов. [18]
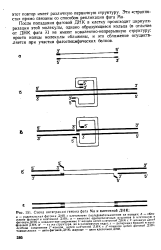 |
Схема интеграции генома фага Ми и клеточной ДНК. [19] |
После попадания фаговой ДНК в клетку происходит циркуля-ризация этой молекулы, однако образующиеся кольца ( в отличие от ДНК фага М не имеют ковалентно-непрерывную структуру: просто концы молекулы сближены, и это сближение осуществляется при участии фагоспецифических белков. [20]
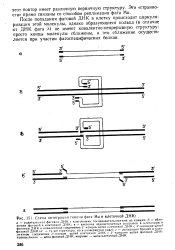 |
Схема интеграции генома фага Ми и клеточной ДНК. [21] |
После попадания фаговой ДНК в клетку происходит циркуля-ризация этой молекулы, однако образующиеся кольца ( в отличие от ДНК фага X) не имеют ковалентно-непрерывную структуру: просто концы молекулы сближены, и это сближение осуществляется при участии фагоспецифических белков. [22]
 |
Схема репликации по способу катящегося кольца. [23] |
К моменту, когда будет завершен полный оборот вокруг минус-цепи, выступающая цепь будет по первичной структуре полностью соответствовать ДНК вириона. На этом этапе при участии белка А происходит отщепление этой цепи с замыканием ее в кольцевую молекулу. Одновременно с этим восстанавливается в первоначальном виде структура RF-II, на которой начитается создание новой плюс-цепи. РНК, можно разделить на две основные группы: обычные РНК-содержащие вирусы и ретровирусы. Последняя группа представляет особый интерес, так как включает в себя вирусы, вызывающие опухолевые заболевания, и вирус HIV, вызывающий синдром приобретенного иммунодефицита, СПИД. [24]
К наглядному представлению о масштабе ожидаемых ускорений приводит рассмотрение механизма реакций, катализируемых а-химотрипси-ном. Если не учитывать специфику реакций, протекающих с участием белков, то можно с достаточной уверенностью считать, что нуклеофилыюсть группы 011 ссрпна в модельных низкомолекулярных соединениях низка, поскольку при физиологических значениях рН 7 - 8 группа ОН слабо ионизована. [25]
 |
Строение ряда типичных представителей вирусов и бактериофагов. [26] |
В заключение этого и двух предыдущих параграфов следует отметить, что сборка многокомпонентных надмолекулярных структур, несмотря на заложенную в их компоненты способность к взаимному узнаванию, скорее всего не является самопроизвольным процессом. В настоящее время накопились факты, указывающие на более сложный, характер образования многосубъединичных белков и нуклеопротеидов, требующих участия вспомогательных белков, которые далее в конечной структуре не присутствуют. Возможно, это относится и к формированию третичной структуры достаточно сложных белков. Установлено, что в этих процессах в большинстве случаев, если не всегда, принимают участие специальные белки, называемые шаперонами. [27]
К числу первых работ в области тройных систем с двумя полимерами следует отнести работу Медведева20, который изучал совместимость ацетата целлюлозы с поливинилацетатом, преследуя цели модификации свойств ацетата целлюлозы. С несколько иными целями был проделан большой цикл экспериментов школой Бунгенберг де йонга21 по изучению системы гуммиарабик - белок - вода и некоторых других систем с участием белков. [28]
Репликация ДНК-геномов требует участия ряда ферментов и дополнительных белков ( см. с. Так, для систем с участием терминального ( концевого) белка такую роль играют сам этот белок и фермент, осуществляющий его нук-леотидилирование. Системы с внутренней инициацией на двухнитевой матрице предполагают участие белков, узнающих оп-сайт. Во всех изученных случаях эти белки закодированы в вирусном геноме. Однако этими ключевыми белками отнюдь не исчерпывается список необходимых участников процесса репликации. Разумеется, обязательно наличие ДНК-полимеразы, а также - в зависимости от схемы репликации и конкретно вирусной системы - набора дополнительных ферментов и белков: праймаз, хеликаз, лигаз, ДНК-связывающих белков и ряда других. В разных вирусных системах эти белки могут иметь либо клеточную природу, либо могут быть закодированы в вирусном геноме. В первом случае все многочисленные компоненты репликационного аппарата, за исключением белка А - это клеточные белки, а во втором - подавляющее их большинство имеет вирусное происхождение. Встречаются случаи, когда фермент, принимающий участие в репликации вирусного генома - это как бы гибридная молекула, построенная из клеточной и вирус-специфической полипептидных цепей. Так, ДНК-полимераза фага Т7 состоит из двух субъединиц - одна из них закодирована в вирусном геноме, а другая представляет собой клеточный белок тиоредоксин. [29]
Репликация ДНК-геномов требует участия ряда ферментов и дополнительных белков ( см. с. Так, для систем с участием терминального ( концевого) белка такую роль играют сам этот белок и фермент, осуществляющий его нук-леотидилирование. Системы с внутренней инициацией на двухнитевой матрице предполагают участие белков, узнающих ori - сайт. Во всех изученных случаях эти белки закодированы в вирусном геноме. Однако этими ключевыми белками отнюдь не исчерпывается список необходимых участников процесса репликации. Разумеется, обязательно наличие ДНК-полимеразы, а также - в зависимости от схемы репликации и конкретно вирусной системы - набора дополнительных ферментов и белков: праймаз, хеликаз, лигаз, ДНК-связывающих белков и ряда других. В разных вирусных системах эти белки могут иметь либо клеточную природу, либо могут быть закодированы в вирусном геноме. В первом случае все многочисленные компоненты репликационного аппарата, за исключением белка А - это клеточные белки, а во втором - подавляющее их большинство имеет вирусное происхождение. Встречаются случаи, когда фермент, принимающий участие в репликации вирусного генома - это как бы гибридная молекула, построенная из клеточной и вирус-специфической полипептидных цепей. Так, ДНК-полимераза фага Т7 состоит из двух субъединиц - одна из них закодирована в вирусном геноме, а другая представляет собой клеточный белок тиоредоксин. [30]
Страницы: 1 2 3