Спиральная участка
Cтраница 2
Интересно, что торцевые и боковые петли часто характеризуются более консервативной первичной структурой, чем спиральные участки. [16]
В плотных образцах с низкой температурой размягчения в качестве типичных элементов макроструктуры были выявлены радиальные, радиально - кольцевые, линейные периодические и спиральные участки. Для образцов с высокой температурой размягчения была характерна пористая губчатая структура с размерами пор от 0.5 до 8 - 10 мм. [17]
Mg достаточно велика и превышает 10 - 4М) указывает на то, что значительная часть оснований находится в регулярных спиральных участках. Образование двойных спиралей РНК на первый взгляд кажется странным. Возможно, что наиболее правдоподобна модель Доти-Фреско, спираль Крика-Уотсона с небольшими петлями, нарушающими регулярную структуру в тех точках, где нет совпадения нужных комплементарных пуриновых и пиримидиновых оснований. В ряде отношений РНК отличается от ДНК по своему поведению в растворах и, следовательно, по своей вторичной и третичной структуре. В ДНК очень слабы полиэлектролитные эффекты, так как это очень прочная спираль и очень жесткая двойная цепь. Соответственно нативная ДНК очень сильно повышает вязкость, а при тепловой денатурации, когда двойная спираль разрушается, вязкость сильно падает. [18]
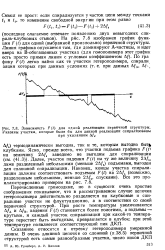 |
Зависимость F ( t для одной реализации первичной структуры. Указаны участки, которые были бы для данной реализации спирализованы. [19] |
Перечисленные громоздкие, но в сущности очень простые соображения показывают, что в рассматриваемом случае цепочка гетерополимера действительно разделяется на клубковые и спиральные участки не флуктуационно, а в соответствии со своей первичной структурой. При росте температуры увеличиваются Д / А и Д / в, падения F ( t) становятся более пологими, а подъемы - крутыми, и каждый участок спирализации при какой-то температуре перестает быть выгодным, превращается в клубок; при этой температуре появляется соответствующий пик ДКП. [20]
Под первичной структурой полифункционального полимера понимается последовательность его мономерных звеньев в цепи, под вторичной структурой - пространственное взаимодействие таких звеньев между собой ( в частности, спиральные участки молекул), под третичной структурой - укладка таких участков в целой молекуле полимера. [21]
Таким образом, из теоретического рассмотрения процесса плавления ДНК вытекает, что с увеличением свободной энергии межплоскостного взаимодействия пар оснований уменьшается ширина интервала перехода и увеличивается число нуклеотидов в спиральных участках в точке перехода. [22]
 |
Модель нуклеиновой кислоты ( по Ф. Крику. [23] |
Третичные структуры образуются за счет определенного расположения в пространстве а-спиралей ДНК или спиральных участков РНК - Конфигурация этих структур меняется в зависимости от различных факторов; так, в растворах с невысокой ионной силой спиральные участки РНК располагаются ( рис. 77 6) перпендикулярно длинной оси частицы РНК, имеющей палочкообразную форму. [24]
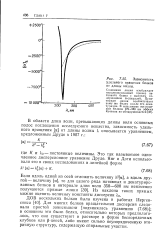 |
Зависимость удельного вращения белков от длины волны. [25] |
Для многих белков вращательная дисперсия следовала простой зависимости [ подчинялась уравнению (7.68) ]; в основном это были белки, относительно которых предполагалось, что они существуют в растворе в форме беспорядочных клубков или р-цепей, либо имеют сильно неупорядоченную вторичную структуру, содержащую спиральные участки. [26]
Как показал анализ, проведенный, в частности, Левиттом 150 ], все известные тРНК могут быть представлены в виде клеверных листов с достаточно большим числом спаренных оснований, так что минорные компоненты ( псевдоуридин, дигидроуридин и др.) всегда оказываются в петлях, а обычные основания составляют спиральные участки. [27]
Эта большая ( для гибкого цепного полимера) величина, если принять модель рис. 8.66, может слагаться, по крайней мере, из трех частей: во-первых, из отрицательной анизотропии одиночных свернутых цепей молекулы ( подобных полистиролу или р-винилнафталину), во-вторых, из незначительной ( вероятно, положительной) анизотропии, вносимой спиральными участками, оси которых ориентированы практически беспорядочно, и, в-третьих, из положительной анизотропии формы. [28]
 |
Модель мс-лекулы РНК ( по Доти. [29] |
Эта большая ( для гибкого цепного полимера) величина, если принять модель рис. 8.66, может слагаться, по крайней мере, из трех частей: во-первых, из отрицательной анизотропии одиночных свернутых цепей молекулы ( подобных полистиролу или ( З - винилнафталину), во-вторых, из незначительной ( вероятно, положительной) анизотропии, вносимой спиральными участками, оси которых ориентированы практически беспорядочно, и, в-третьих, из положительной анизотропии формы. [30]
Страницы: 1 2 3 4