Спиральная участка
Cтраница 4
Оптическая активность белков, полисахаридов и нуклеиновых кислот обусловлена их оптически активными компонентами - аминокислотами и сахарами, а также асимметрией их вторичной структуры, имеющей форму право - или левовинтовых спиралей. Денатурированный белок имеет конформацию беспорядочного клубка и поэтому дает оптическое вращение, отличное от того, которое дает соответствующий нативный белок, содержащий спиральные участки. Оптическое вращение растворов амилопектина, имеющего в основном неспиральное строение, отличается от оптического вращения свежеприготовленной спиральной амилозы, если проводить сравнение в пересчете на один и тот же вес глюкозы. Изменения во вторичной структуре макромолекул удается регистрировать путем измерения удельного вращения не только по всему спектру, но и при одной длине волны. Уже с давних пор известно, что белок по мере денатурации приобретает все более и более отрицательное удельное вращение. Величины [ а ] о для полностью денатурированных белков и беспорядочно свернутых полипептидов лежат в интервале от - 90 до - 125, тогда как удельное вращение белков в нативном состоянии составляет 100 и больше. Изменения конформации белков, обусловленные изменением рН, также отражаются на величине удельного вращения. Все эти свойства белковых растворов известны по наблюдениям их удельного вращения при одной длине волны - как правило, при длине волны D-линии натрия. [46]
 |
Конформационные карты трипептида Ала-Ала, построенные минимизацией по ф2, №, минимум энергии второго остатка соответствует конформации В. W7A ф240 15. г 2330 5. Ш1Ф250 15. г 52330 5с. [47] |
Такая упрощенная модель, по утверждению Котельчука и Шера-га [21] дает более 75 % правильных предсказании конформации пептидных единиц в четырех белках. Однако слабая ко-оперативность вдоль цепи, связанная с взаимодействием пептидных единиц, все же имеется и она резко увеличивает трудности предсказаний, покуда детально не изучены конформации различных комбинаций остатков и их относительные стабильности в трипептидных фрагментах или в относительно больших спиральных участках. В самом деле, вместо того, чтобы начинать поиск минимума с однозначно или почти однозначно определяемого нулевого приближения, приходится считаться со слабой кооперативностью вдоль цепи. [48]
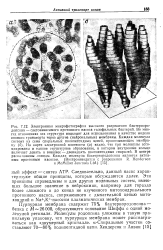
Пурпурная мембрана содержит 75 % бактериородопсина - белка с М-26000, образующего основание Шиффа с одной молекулой ретиналя. Молекулы родопсина уложены в такую правильную упаковку, что пурпурная мембрана может рассматриваться как двумерный кристалл. Спиральные участки составляют 70 - 80 % полипептидной цепи. [50]
Высокая степень гидрофобности центрального участка реакционного центра свидетельствует о том, что пять а-спиральных сегментов субъединиц Л и С, а также N-конце во и фрагмент субъединицы Т расположены а толще мембраны. Все эти I I участков и составляют мембранную часть комплекса. Спиральные участки расположены в пространстве практически перпендикулярно плоскости мембраны. Субъединица Т и цитохром с связываются с внутренней и наружной поверхностями мембраны соответственно, причем цитохром с не содержит участков, пронизывающих мембрану. По два мембранных фрагмента от каждой из субъединиц Л и С участвуют в связывании негемового железа. Более того, четыре лиганда, необходимые для связывания атома железа, обеспечиваются этими же четырьмя спиралями. Лиганды для ионов магния молекул бактериохлоро-филла - остатки гистидина - расположены у самого входа ct - спи-ральных сегментов в мембрану. Что же касается негемового железа, то в его связывании участвуют четыре остатка гистидина субъединиц Л и С и глутаминовая кислота субъединицы С. [51]
Как связаны те закономерности, которые мы рассматривали с пространственной структурой глобулярных белков. Как мы уже отмечали, Брант и Шиммель [91], Рамачандран и Саси-секхаран [19], а также Шерага [20] рассматривали распределение точек, соответствующих углам р и ty в этих белках, на конформационных картах дипептидов. Очевидно, спиральные участки белков не представляют интереса ( равно как и области - структуры в лизоциме), их углы ф и - ф находятся вблизи минимумов конформационной карты и потому мы приводим на рис. 26 только точки, относящиеся к неспиральным участкам. [52]
Рибосомная РНК ( рРНК) находится в комплексе с белками в рибосомах. На ее долю приходится до 65 процентов от всей РНК. Вторичная структура представлена спиральными участками, соединенными изогнутой одиночной цепью. Третичная структура рРНК - скелет рибосомы, имеет форму палочки или клубка; снаружи находятся рибосомальные белки. [53]
Оценка степени спирализации по величине оптической активности также весьма условна. Это говорит о том, что фрагмент трипсина состоит преимущественно из левых спиралей, тогда как в макромолекуле трипсина преобладают в целом правовращающие структуры. Отсюда ясно, что спиральные участки в трипсине могут давать слагаемые разного знака, и величина оптической активности оказывается по абсолютной величине меньшей, чем в случае белка с той же степенью спирализации, но с одними правыми спиралями. Аналогичные правые ( цепь В) и левые ( цепь А) спирали были обнаружены и в молекуле инсулина. [54]
Очевидно, при охлаждении часть спиральных участков восстанавливается. Однако вязкость раствора после охлаждения не достигает тех высоких значений, какие она имела до нагревания. Это свидетельствует о том, что восстановленные спиральные участки имеют небольшую длину. Повторное нагревание охлажденного раствора показывает, что переход теперь происходит в широком температурном интервале. Картина, наблюдаемая при повторном нагревании, воспроизводится при последующих циклах нагревания и охлаждения. Наличие широкой области перехода указывает на то, что в растворе образовались короткие спиральные участки различной длины и состава за счет неправильного спаривания оснований, происходящего, вероятно, в пределах одной полинуклеотидной цепочки. Нагревание до температур, лежащих выше области перехода, приводит к разделению цепей. Охлаждение, следующее за нагреванием, приводит лишь к тому, что основания, принадлежащие одной цепи, неспецифически взаимодействуют, образуя водородные связи друг с другом и с основаниями других цепей. [55]
Эти свойства напоминают свойства молекул в форме достаточно жестких стержней, обладающих сильным отрицательным двулучепреломлением. Отсюда следует, что в тРНК основания должны быть ориентированы в основном перпендикулярно к длинной оси молекулы. Сегментная анизотропия тРНК позволяет предсказать, что спиральные участки и молекулы состоят из 9 - 10 пар оснований. [56]
Здесь же следует упомянуть работу Миллера [5] по ферментативному ращеплению поли-а-1 - глутаминовой кислоты. Как известно, относительно неспецифический фермент папаин гидро-лизует поли-1 - глутаминовую кислоту. Оказалось, что фермент действует только на спиральные участки цепи и что все пептидные связи в равной степени чувствительны к действию фермента, за исключением двух связей, ближайших к концу цепи. [57]
Страницы: 1 2 3 4