-
Cтраница 1
Фаг прикрепляется к бактериальной клетке, во много раз превосходящей его по размерам, с помощью хвостового отростка. Эксперименты с двойной меткой показывают, что ДНК, меченная Р32, впрыскивается в бактерию, а белок, содержащий S35, остается снаружи. Сразу же после такой инъекции синтез ДНК, РНК и белков клетки-хозяина прекращается. В дальнейшем всеми синтетическими механизмами клетки управляет ДНК фага. Через 2 мин появляется фагоспецифич-ная РНК; через 4 мин можно обнаружить новый белок; через б мин после инъекции начинается синтез фаговой ДНК, причем он идет примерно в 5 раз быстрее, чем синтез ДНК хозяина в незараженных клетках. Через 20 мин образуется примерно 100 новых фаговых частиц, которые освобождаются из клетки. [1]
 |
Гомологичная рекомбинация между двумя кольцевыми репликонами.| Механизм мономери. [2] |
Фаг Р1 эффективно обходит эту сложность за счет кодируемой им системы сайт-специфической рекомбинации. Продукт фагового гена ere с высокой частотой проводит рекомбинацию между двумя специальными последовательностями loxP, находящимися на одной молекуле ДНК. In vitro очищенный Сге-белок может осуществлять всю реакцию без каких-либо дополнительных факторов и без доноров энергии. Как н другие белки сайт-специфической рекомбинации, он напоминает топоизомеразы в том отношении, что в ходе рекомбинации вносит разрывы в ДНК и ковалентно связывается с одним из образовавшихся концов, запасая тем самым энергию разорванной связи. [3]
Фаг прикрепляется к бактерии хвостовым отростком, через который пропускает содержимое фаговой частицы. ДНК фага меняет обмен веществ бактерии, направляя его по новому пути. В этом процессе она и спользует ферменты бактерий. [4]
Фаги размножаются быстрее, чем бактерии. Это относительно легко показать, если во взвесь бактерий внести возможно большее число фаговых частиц: первоначально мутная ( из-за содержащихся в ней бактериальных клеток) суспензия очень скоро становится совершенно прозрачной, так как все бактерии лизируются. [5]
Фаг весьма редко переносит одновременно два признака; в некоторых случаях это даже совершенно исключено, что, впрочем, легко понять. Подсчитано, что бактериальный геном примерно в 100 раз длиннее, чем геном фага; поэтому совместно могут передаваться разве что те гены, которые удалены друг от друга не более чем на одну сотую длины генома. [6]
Фаги Т2 и Т4 способны давать большое число двойных и даже заметное число тройных рекомбинантов. [7]
Фаг, подходя к микробу, присасывается к нему горлышком. На конце горлышка находятся белки-ферменты, делающие оболочку микроба проницаемой. Как только фермент сделал свое дело, ДНК фага как бы впрыскивается внутрь микроба, а у стенки остается пустая оболочка-бутылочка, или, как ее называют, тень фага. ДНК, попавшая в микроб, довольно быстро исчезает, во всяком случае обнаружить ее не удается. Но примерно уже через 30 минут клетка микроба оказывается набитой множеством фагов. Фактически микроб исчез, превратился в мешок с фагами. [8]
Фаг прикрепляется к бактерии хвостовым отростком, через который выпускает содержимое фаговой частицы. ДНК фага меняет обмен веществ бактерии, направляя его по новому пути. В этом процессе она использует ферменты бактерий. [9]
Фаг - -, угол установки 4 лопаток диффузора при выходе, соотношение диаметров рабочего колеса Я, - р, диффузора - - и некоторыми другими. [10]
Фаги, как и другие вирусы, обладают антигенными свойствами и содержат группоспецифические и типоспецифические антигены. [11]
Фаги могут применяться в качестве диагностических препаратов для установления рода и вида бактерий, выделенных в ходе бактериологического исследования. Однако чаще их применяют для лечения и профилактики некоторых инфекционных заболеваний. [12]
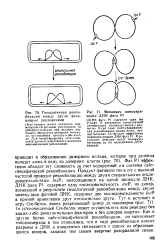
Фаг Р1 эффективно обходит эту сложность за счет кодируемой им системы сайт-специфической рекомбинации. Продукт фагового гена ere с высокой частотой проводит рекомбинацию между двумя специальными последовательностями loxP, находящимися на одной молекуле ДНК. ДНК фага Р1 содержит одну последовательность loxP, но димер, возникший в результате гомологичной рекомбинации между двумя молекулами фаговой ДНК, содержит две последовательности / о. Сге-белок может осуществлять всю реакцию без каких-либо дополнительных факторов и без доноров энергии Как и другие белки сайт-специфической рекомбинации, он напоминает топоизомеразы в том отношении, что в ходе рекомбинации вносит разрывы в ДНК и ковалентно связывается с одним из образовавшихся концов, запасая тем самым энергию разорванной связи. [14]
Фаги ( лямбдоидные фаги) обычно встраиваются в одно место хромосомы, но при определенных условиях могут располагаться и в др. участках генома. Фаги способны включаться в любые места бактериальной хромосомы, а также в ДНК мн. Интеграция лямбдо-идных фагов обеспечивается ферментной системой, состоящей из клеточных белков и белков, кодируемых геномом фага. [15]
Страницы: 1 2 3 4 5