Фактор - элонгация
Cтраница 1
Фактор элонгации Ти GDP по форме напоминает головастика. [1]
Фактор элонгации Ти GDP по форме напоминает головастика. Форма молекулы заметно отличается от формы обычных глобулярных белков. Это - головастик, состоящий из глобулярного домена ( головы) и удлиненного тонкого домена ( хвоста), разделенных расщелиной с меньшей плотностью. Голова содержит несколько а-спиралей и, по-видимому, имеет довольно жесткую структуру. Хвост, напротив, более лабилен и, по-видимому, представляет собой р-структуру. Интересно, что голова и хвост имеют и второе сочленение, так что в результате возникает некая циклическая структура. Вероятно, тРНК присоединяется вблизи отверстия этого цикла в большом желобе между доменами. Хорошо видно относительнее смещение доменов во время стадии элонгации на рибосоме. В L7 / L12 подвижный домен находится со стороны N-конца. [2]
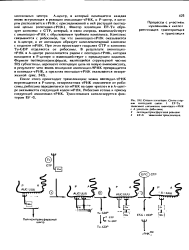 |
Стадии элонгации. Синтез первой пептидной связи. / EF-Tu зависимое связывание аминоацнч-т РНК с А участком рибосомы. [3] |
Фактор элонгации EF-Tu образует комплекс с GTP, который, в свою очередь, взаимодействует с аминоацил-т РНК с образованием тройного комплекса. При этом происходит гидролиз GTP и комплекс Tu-GDP отделяется от рибосомы. В результате аминоацил-т РНК в А-центре располагается рядом с пептидил-т РНК, которая находится в Р - центре и взаимодействует с предыдущим кодоном. [4]
Фактор элонгации Ти GDP по форме напоминает головастика. Форма молекулы заметно отличается от формы обычных глобулярных белков. Это - головастик, состоящий из глобулярного домена ( головы) и удлиненного тонкого домена ( хвоста), разделенных расщелиной с меньшей плотностью. Голова содержит несколько а-спиралей и, по-видимому, имеет довольно жесткую структуру. Хвост, напротив, более лабилен и, по-видимому, представляет собой р-структуру. Интересно, что голова и хвост имеют и второе сочленение, так что в результате возникает некая циклическая структура. Вероятно, тРНК присоединяется вблизи отверстия этого цикла в большом желобе между доменами. Хорошо видно относительнее смещение доменов во время стадии элонгации на рибосоме. В L7 / L12 подвижный домен находится со стороны N-конца. [5]
Эукариотический фактор элонгации EF-2 утрачивает неспецифическое сродство к РНК в результате АДФ-рибозилирования. [6]
 |
Генетически сконструированный химерный комплекс CD4 - экзотоксин A Pseudomonas. Использован промотор бактериофага Т7 Е. coli. [7] |
Затем экзотокси-новая часть химерного белка инактивирует фактор элонгации EF-2, участвующий в синтезе белка. Это препятствует дальнейшему синтезу белка, что в конце концов приводит к гибели клетки. [8]
Транслокация катализируется довольно крупным белком, называемым фактором элонгации G ( EF-G) у прокариот или фактором элонгации 2 ( EF-2) у эукариот. EF-G ( или, соответственно, EF-2) взаимодействует с ГТФ и с рибосомой. При этом взаимодействии наводится ГТФазная активность, и ГТФ расщепляется до ГДФ и ортофосфата. При взаимодействии ( комплексообразовании) EF-G и ГТФ с претранслока-ционной рибосомой происходит быстрая транслокация, a EF-G, ГДФ и ортофосфат освобождаются из комплекса с рибосомой. [9]
Функционирование факторов терминации, так же как факторов элонгации и одного из факторов инициации, зависит от ГТФ. Для эукарио-тических систем это показано достаточно четко: ГТФ, так же как и его нерасщепляемый аналог, стимулирует кодонзависимое связывание RF с рибосомой, с другой стороны, RF на рибосоме обладает ГТФазной активностью, которая, видимо, нужна для расщепления ГТФ с целью освобождения RF с терминировавшей рибосомы. [10]
Элонгация осуществляется при помощи белков цитозоля, называемых факторами элонгации. [11]
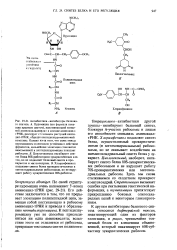
К другим ингибиторам белкового синтеза относятся дифтерийный токсин, инактивирующий один из факторов элонгации, и рицин, чрезвычайно токсичный белок из клещевины обыкновенной, который инактивирует 608-суб-частицу эукариотических рибосом. [13]
ДНК, РНК и рибосом и, входя в состав аминоацил-т РНК-синтетаз и фактора элонгации белковой цепи у млекопитающих ( EF-1), играет важную роль в процессе трансляции и незаменим, таким образом, на многих ключевых этапах экспрессии гена. В этой связи становится понятной задержка роста и развития у человека и лабораторных животных при алиментарной или наследственной недостаточности цннка. Необходимость цинка для нормального протекания всех этапов клеточного цикла и для функционирования генетического аппарата продемонстрирована на примере эвглены [ Vallee В. [14]
В частности, возможные изменения в популяции изоакцепторных тРНК, а также изменения в концентрациях факторов элонгации и аминоацил-т РНК-синтетаз могут влиять на избирательную регуляцию элонгации. [15]
Страницы: 1 2 3