Фактор - элонгация
Cтраница 2
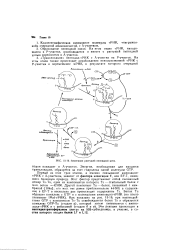 |
Элонгация растущей пептидной цепи. [16] |
Первый из этих трех этапов, а именно связывание аминоацил-т РНК с А-участком, зависит от фактора элонгации Т, или EF-T, имеющего белковую природу. Этот фактор представляет собой смешанный димер Ts-Tu, один из компонентов которого Ts - стабильный белок с мол. Другой компонент Ти - белок, связанный с мембраной [ 104а ]; его мол. EF-T в несколько раз превосходит содержание Ts. [17]
Попеременно меняя концентрацию Mg2 в бесклеточной системе, можно было бы в известной мере имитировать действие факторов элонгации и тем самым значительно ускорить бесфакторный элонгационный цикл. [18]
Транслокация катализируется довольно крупным белком, называемым фактором элонгации G ( EF-G) у прокариот или фактором элонгации 2 ( EF-2) у эукариот. EF-G ( или, соответственно, EF-2) взаимодействует с ГТФ и с рибосомой. При этом взаимодействии наводится ГТФазная активность, и ГТФ расщепляется до ГДФ и ортофосфата. При взаимодействии ( комплексообразовании) EF-G и ГТФ с претранслока-ционной рибосомой происходит быстрая транслокация, a EF-G, ГДФ и ортофосфат освобождаются из комплекса с рибосомой. [19]
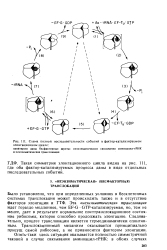 |
Схема полной последовательности событий в фактор-катализируемом элонгационном цикле. [20] |
Было установлено, что при определенных условиях в бесклеточных системах транслокация может происходить также и в отсутствие факторов элонгации и ГТФ. Эта неэнзиматическая транслокация идет гораздо медленнее, чем EF-G GTP-катализируемая, но, тем не менее, дает в результате нормальное посттранслокационное состояние рибосомы, которое способно продолжать элонгацию. Следовательно, процесс транслокации является термодинамически спонтанным. Транслокационный механизм оказывается принципиально присущ самой рибосоме, а не привносится фактором элонгации. [21]
 |
Расположение функциональных центров. на субчастицах рибосомы ( по А. С. Спирину, 1999. [22] |
Цепь мРНК связана с малой субчастицей между участками Р и А. Пеп-тидилтрансферазный центр РТС и факторы элонгации EF располагаются на большой субчастице. [23]
Таким образом, наши представления о взаимодействиях между белками и нуклеиновыми кислотами пока еще очень ограниченны. Примеры рибосомальных белков и фактора элонгации Ти GDP показывают, что некоторые нуклео-протеиды имеют весьма своеобразную структуру. [24]
Весьма интересен молекулярный механизм действия дифтерийного токсина. Он оказался наделенным способностью катализировать реакцию АДФ-рибозилирования фактора элонгации эукариот ( eEF - 2), выключая тем самым его из участия в синтезе белка. Резистентность многих животных к дифтерийному токсину, вероятнее всего, обусловлена трудностью или полным отсутствием проникновения ( транспорта) токсина через мембрану клеток. [25]
Она катализируется крупным мономерным белком, обозначаемым как фактор элонгации О ( ЕР-О) у прокариот или еЕР - 2 у эукариот, с использованием молекулы ГТФ. [26]
Все нативные 40S частицы обязательно содержат eIF - З в эквимолярном количестве. Нативные 60S субчастицы могут иметь, по-видимому, связанные факторы элонгации. [27]
Элонгация представляет собой образование и удлинение полипептидной цепи, формирующейся на рибосоме. Этот процесс проходит при участии ГТФ и трех факторов элонгации. Эти факторы у прокариот имеют обозначения: EF-TU, EF-TS и EF-G, или просто: Tu, Ts и G. Фактор элонгации Ти образует комплекс с ГТФ, который связывается со всеми аминоацил-т РНК в цитоплазме. Ти обладает ГТФ-азной активностью и гидролизует ГТФ. После этого Ти, ГДФ и Рг удаляются из рибосомы и исходный комплекс регенерирует при помощи Ts и ГТФ. [28]
Имеется, однако, одно свойство EF-2 ( не имеющее прямого отношения к промотированию транслокации), которое полностью утрачивается после АДФ-рибозилирования. Это - неспецифическая РНК-связывающая способность, которая свойственна эукариотическим факторам элонгации ( так же как и аминоацил-т РНК - синтетезам - см. A. Благодаря этой способности значительная часть EF-2 ( так же как и EF-1) эукариотической клетки оказывается компартментализованной вокруг полирибосом за счет лабильной и обратимой ассоциации с их. АДФ-рибозилирование EF-2 приводит к исчезновению неспецифической РНК-связывающей способности фактора и к его полной декомпартментализации из полири-босомных структур. Может быть, что этот эффект, приводящий к резкому уменьшению локальной концентрации фактора вблизи полирибосом, ответствен, по крайней мере частично, за ингибирование белкового синтеза под действием А-фрагмента дифтерийного токсина. [29]
Процесс элонгации принято делить на 3 стадии: узнавание кодона и связывание аминоацил-т РНК, образование пептидной связи и транслокация. На I стадии в соответствии с природой кодона мРНК в свободный А-участок рибосомы доставляется аминоацил-т РНК при участии фактора элонгации Tu. Этот процесс требует затраты энергии и сопряжен с гидролизом ГТФ и образованием прочно связанного комплекса Tu-ГТФ. Образовавшийся комплекс подвергается диссоциации только в присутствии второго фактора элонгации Ts, при котором освободившийся фактор Tu может вновь, соединяясь с молекулой ГТФ, принять участие в доставке аа-т РНК в рибосому. С этого момента начинается II стадия элонгации-образование первой пептидной связи. Для этого в рибосоме осуществляется ферментативная реакция транспептидирования между формилметионил-т РНК в П - центре и новой аа-т РНК в А-центре. В процессе этой реакции остаток формилметионина переносится на свободную NH2 - rpynny аа-т РНК и замыкается первая пептидная связь в будущей полипептидной цепи. Параллельно из пеп-тидильного центра освобождается тРНК Мет в цитозоль. Фермент, катализирующий реакцию транспептирования, получил название пептидил-трансферазы ( рис. 14.7); он, вероятнее всего, является составной частью белков 50S субчастицы. [30]
Страницы: 1 2 3