Фосфорилаза
Cтраница 1
Фосфорилаза а представляет собой димер, состоящий из двух идентичных субъединиц, в каждой из которых имеется один специфический остаток се-рина, фосфорилированный по гидрок-сильной группе. Эти остатки фосфосери-на необходимы для максимальной активности фермента. Фосфатные группы, соединенные с остатками серина, можно удалить из фосфорилазы а с помощью фермента, называемого фосфа-тазой фосфорилазы, который катализирует гидролитический разрыв связи между фосфатом и остатком серина. [1]
Фосфорилаза b может снова реактивироваться, т.е. превратиться в активную фосфорилазу а. [2]

Фосфорилаза а была получена в кристаллическом виде ( мол. Взаимопревращения двух этих форм гликоген-фосфорилазы происходят под действием специфичных ферментов, катализирующих процесс ковалентной модификации ( разд. [4]
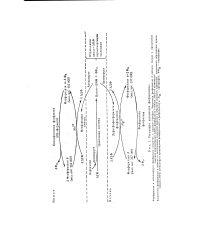 |
Регуляция активности фосфорилазы. [5] |
Фосфорилаза а катализирует фосфоролиз гликогена в глюкозо-1 - фосфат. [6]
 |
Превращения фосфорилазы а в фосфорилазу 7 в и обратная реакция. [7] |
Фосфорилаза отщепляет глюкозные остатки в виде глюкозо-1 - фосфата, начиная от периферического конца наружных ветвей. При приближении к х - 1 6-свя-зям ее действие прекращается. Олиготрансфераза перестраивает оставшиеся культи, обнажая глюкозный остаток, соединенный а-1 6-связью. Только на такой обнаженный остаток может действовать ами-ло-1 6-глюкозидаза. После удаления этого остатка вновь открывается поле деятельности для фосфорилазы. [8]
Фосфорилаза и гликогенсинтаза имеют субъединичное строение. Неактивная ( малоактивная) фосфорилаза b состоит из нефосфори-лированных димеров и под действием фермента киназы фосфорилазы с участием АТФ переходит в фосфорилазу а ( активная форма), которая представлена фосфорилированными тетрамерами. Дефосфори-лирование фермента, т.е. переход формы a - b осуществляется с помощью фермента протеинфосфатазы. Гликогенсинтаза, наоборот, в неактивном состоянии ( D-форма) является фосфорилированным тетрамером. Переход в активную I-форму ( дефосфорилированный тетрамер) осуществляется под действием протеинфосфатазы. [9]
Фосфорилаза ( Ь) неактивна ( по крайней мере в отсутствие AMP - см. ниже) и существует в виде димера. Обратная реакция катализируется отдельным ферментом - фосфатазой фос-форидазы. [10]
Фосфорилаза ( а) агрегирует далее и образует тетрамер. Эта стадия усиления ( амплификации:) очень напоминает эффекты электронных цепей. [11]
Фосфорилаза крахмала обнаружена во многих растениях, а из картофеля выделена в кристаллическом виде. Этот фермент катализирует обратимый перенос глюкозильных остатков между глюкозо-1 - фосфатом и невосстанавливающим концом глюкозной цепочки, содержащей сс-1 4-связи. В качестве акцепторов могут действовать различные соединения; однако для фермента, по-видимому, необходима невосстанавливающая цепочка, оканчивающаяся по меньшей мере тремя глюкозильными остатками, соединенными а-1 4-связями, причем замещение в остатках может происходить только у 1-го и 4-го атомов углерода. Наименьшей молекулой, способной действовать в качестве акцептора, является молекула мальтотриозы. Чем больше молекулы, тем они эффективнее; наиболее эффективными оказались соединения с цепочкой, состоящей примерно из 20 глюкозильных остатков, соединенных а-1 4-свя-зями. Фермент действует более активно на разветвленный акцептор, содержащий несколько таких цепочек, чем на акцептор с неразветвленной цепью. Наибольшую активность фермент проявляет по отношению к амилопектину. Фермент не изменяет число невосстанавливающих концевых групп; небольшая молекула акцептора может послужить исходной для образования молекулы больших размеров. Поэтому такой акцептор ( в данной реакции и других ей подобных) называют затравкой. Однако он представляет собой просто второй реагент бимолекулярной реакции. [12]
Неактивная фосфорилаза ( Ь) превращается в активную фосфорилазу ( а) в результате ее фосфорилирования фосфорилаза-6 - киназой. Однако последняя также существует в фосфорилированной ( активной) и нефосфорилированной ( неактивной) формах. Протеинкиназа может фосфорилировать не только фосфо-рилазокиназу, но также многие другие белки, такие как казеин и даже гликогенсинтетазу. Фосфорилирование гликогенсинтетазы инактивирует фермент, и в итоге сАМР не только активирует деградацию гликогена, но также и выключает его синтез. В результате каскадного процесса, описанного выше, эффект малого количества сАМР усиливается, сАМР может осуществлять очень тонкий контроль за процессом деградации гликогена. [13]
 |
Схема строения комплекса амилозы с иодом. [14] |
Действие фосфорилазы обратимо, причем вначале было открыто ее расщепляющее действие-фосфоролиз, в связи с этим возникло название этого фермента. [15]
Страницы: 1 2 3 4 5