Фосфорилирование - белок
Cтраница 2
Процесс дефосфорилирования белков происходит под действием ферментов группы фосфопротеинфосфатаз. Фосфорилирование белков цАМФ - зави-симыми протеинкиназами не ограничивается цитоплазмой. С-Каталитиче - ские субъединицы протеинкиназ способны пересекать ядерные мембраны и, фосфорилируя ядерные белки - гистоны, регулировать генную активность клеток. [16]
Фосфорилирование сопровождается активацией или инактивацией ферментов, напр, гликозилтрансфераэ, а также изменением физ. Обратимое фосфорилирование белков контролирует, напр. [17]
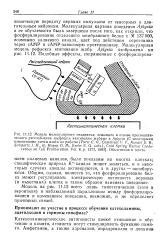
К - канала может меняться, и в некоторых случаях каналы активируются, а в других - ингибиру-ются. Общим, однако, является то, что фосфорилирование белка может регулировать электрическую активность нервных ( и мышечных) клеток и таким образом изменять поведение. [19]
В то время как механизм пассивного транспорта, как правило, известен, механизм сопряжения транспорта вещества с обеспечивающей этот процесс энергией химической реакцией остается не ясным. Вероятно, при функционировании Ма К - АТРазы происходит временное фосфорилирование белка переходящими с АТР-ионами фосфата. [20]
 |
Ковалентная регуляция гликогенфосфорилазы. [21] |
Активность многих ферментов регулируется цАМФ - зависимым фосфо-рилированием, соответственно большинство гормонов белково-пептидной природы активирует этот процесс. Однако ряд гормонов оказывает тормозящий эффект на аденилатциклазу, соответственно снижая уровень цАМФ и фосфорилирование белков. В частности, гормон соматостатин, соединяясь со своим специфическим рецептором-ингибиторным G-белком ( Gj, являющимся структурным гомологом Оз-белка ( см. ранее), ингиби-рует аденилатциклазу и синтез цАМФ, т.е. вызывает эффект, прямо противоположный вызываемому адреналином и глюкагоном. [22]
Синтез и секреция Т3 и Т4 контролируются тиролиберином и тиротропином. Тиротропин оказывает следующие эффекты: стимулирует активный транспорт 1 - против 500-кратного градиента в полость фолликула за счет цАМФ - зависимого фосфорилирования белков клеточных мембран; усиливает транскрипцию и трансляцию тироглобулина; стимулирует рост эпителиальных клеток, формирующих фолликулы, а в фолликулярном коллоиде - иодирование тирозилов; по аденилатциклазному механизму стимулирует синтез Т3, Т4 ( аналогично действуют адреналин и PGE2); стимулирует секрецию иодированного тироглобулина путем пиноцитоза и отщепления Т3 и Т4 при слиянии пиноцитозных пузырьков с мембранами лизосом ( протеолитическим путем), а также поступление Т3 и Т4 в кровь и лимфу. [23]
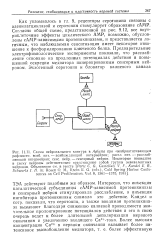
Согласно общей схеме, представленной на рис. 9.12, все внутриклеточные эффекты циклического AMP, возможно, обусловлены сАМР - зависимыми протеинкиназами, и представляется логичным, что наблюдаемая сенситизация имеет некоторое отношение к фосфорилированию ключевого белка. Предварительные электрофизиологические эксперименты показали, что расслабление основано на продленных потенциалах действия и повышенной секреции медиатора иннервированным сенсорным нейроном. [25]
Окситоцин оказывает стимулирующее действие на гладкую мускулатуру матки, а также способствует сокращению миоэпи-телиальных клеток в районе альвеол молочной железы. Расслабляюще действует на гладкие мышцы сосудов, вызывая временную артериальную гипотонию. Молекулярные механизмы эффектов обусловлены генерированием цАМФ и фосфорилированием соответствующих белков. [26]
Эта реакция ведет к совершению механической работы и к частичному переходу энергии в теплоту. Первая реакция пока в точности неизвестна. Однако имеются веские причины считать, что это - реакция переноса эфирносвязанной фосфатной группы с АТФ на белок, или реакция фосфорилирования белка. [27]
Про те инки на за - это внутриклеточный фермент, через который цАМФ реализует свой эффект. Протеинкиназа может существовать в 2 формах. В отсутствие цАМФ Протеинкиназа представлена в виде тетрамерного комплекса, состоящего из двух каталитических ( С2) и двух регуляторных ( Rj) субъединиц с мол. В присутствии цАМФ протеинкиназный комплекс обратимо диссоциирует на одну К2 - субъединицу и две свободные каталитические субъединицы С; последние обладают ферментативной активностью, катализируя фосфорилирование белков и ферментов, соответственно изменяя клеточную активность. [28]
Катехоламины действуют на клетки-мишени по мембрано-опосредованному механизму, чему в немалой степени способствует гидроксилирование кольца и боковой цепи этих соединений. Катехоламины взаимодействуют с а - и р-адренергическими рецепторами, локализованными в мембранах клеток-мишеней. Адреналин взаимодействует с обоими типами рецепторов, а норадреналин преимущественно с а-рецепторами. Группа ах -, а2 - рецепторов проявляет эффекты сосудосуживающего действия, сокращения гладких мышц, ингибирования липолиза. Действие ( 3-рецепторов связано с активацией аденилатциклазы, образованием цАМФ и последующим фосфорилированием белков. Например, адреналин, взаимодействуя с р-рецепторами через систему вторичных посредников, активирует протеинкиназу, которая фосфорилирует ряд цитоплазматических белков. Таким образом, адреналин регулирует гликогенолиз в печени и в мышцах, а также глюконеогенез в печени. [29]
Некоторые белки, модифицируемые ковалентным фосфорилиро-ванием, имеют типичные свойства, общие с нуклеотидсвязываю-щими белками. Обычно фосфор ил ируется боковая цепь остатков Ser, Thr или His. Часто перед таким остатком или сразу за ним располагается остаток Gly; в большинстве случаев остаток, находящийся в цепи на две позиции дальше, оказывается положительно заряженным Lys или Arg. Гипотеза о том, что обратимое фосфорилирование данного остатка управляет конформационными изменениями фосфопро-теида [175, 693], хорошо коррелирует с тем фактом, что в лактат-дегидрогеназе [232] и в аденилаткиназе [688] петля является эпицентром больших конформационных изменений. Таким образом, исследования нуклеотидсвязывающих белков могут способствовать прояснению статических и динамических аспектов фосфорилирования белков. [30]
Страницы: 1 2 3