Фосфофруктокиназа
Cтраница 2
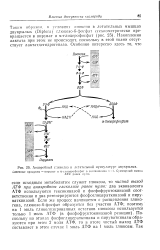 |
Анаэробный гликолиз в летательной мускулатуре двукрылых. [16] |
АТФ при анаэробном гликолизе равен нулю: два эквивалента АТФ используются гексокиназой и фосфофруктокиназой соответственно и два регенерируются фосфоглицераткиназои и пиру-ваткиназой. Поскольку на этапах фосфоглицераткиназы и пируваткиназы образуются при этом 2 моля АТФ, то в итоге чистый выход АТФ составляет в этом случае 1 моль АТФ на 1 моль глюкозы. [17]
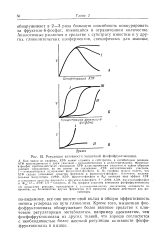
Кроме того, мышечная фос-фофруктокиназа обнаруживает более высокое сродство к ключевым регуляторным метаболитам, например аденилатам, чем фосфофруктокиназы из других тканей, что хорошо согласуется с необходимостью более жесткой регуляции активности фосфофруктокиназы в мышце. [19]
Таким образом, сопряжение реакции, катализируемой фосфофруктокиназой, с реакцией, катализируемой а-глицерофосфат-дегидрогеназой, позволяет на основе данных, полученных при регистрации оптической плотности при 340 нм, судить о скорости реакции, катализируемой фосфофруктокиназой. [20]
Предшествующие промежуточные продукты могут использоваться в других реакциях, и поэтому стадия д представляет собой важную точку регуляции. Фосфофруктокиназа ингибируется высокими концентрациями АТФ и активируется АДФ и АМФ; гликолиз протекает интенсивнее при низком отношении АТФ: АДФ. Биологический смысл этого таков, что, как видно из общего уравнения гликолиза ( стр. АТФ и при понижении концентрации АТФ в клетке гликолиз усиливается. [21]
Ферменты гликолитического превращения глюкозы - фосфофруктокиназа и фосфоглюко-мутаза - были выделены японскими исследователями из клеток экстремально-термофильной бактерии Flavobacterium thermophilus НВ-8. Так, фосфофруктокиназа не инактивировалась в течение 1 ч при 80 С, и за то же время потеря активности составила всего 10 % при температуре прогрева, равной 90 С. [22]
В связи с этим фосфофруктокиназа имеет два центра связывания АТФ - один для АТФ как субстрата, другой - для АТФ как аллостерического ингибитора. Это означает, что скорость образования фруктозо-1 б-дифосфата как функция концентрации АТФ проходит через максимум. Наоборот, АМФ и АДФ, высокий уровень которых является сигналом о дефиците энергии, являются аллостерическими активаторами фермента. [23]
Находится ли данная физиологическая реакция в состоянии равновесия. Что говорит этот эксперимент о роли фосфофруктокиназы в качестве регулятор-ного фермента. [24]
Вторым регуляторным пунктом глю-конеогенеза служит реакция, катализируемая фруктозодифосфатазой, ферментом, на который резкое ингибирующее действие оказывает AMP. Так как соответствующий фермент гликолитиче-ского пути, фосфофруктокиназа, активируется AMP и ADP, а ингибируется цитратом и АТР ( разд. [25]
Гли-коген-фосфорилаза, катализирующая превращение глюкозных единиц гликогена в глюкозо-1 - фосфат, принадлежит к числу регуляторных ферментов и существует в двух формах: более активной ( фосфорилаза а) и менее активной ( фос-форилаза Ъ); стимулирующее действие на фосфорилазу b оказывает AMP. Роль главного регуляторного фермента в последовательности реакций гликолиза играет фосфофруктокиназа, которую ин-гибируют АТР и цитрат и стимулирует AMP. Вторым регуляторным пунктом гликолиза является пируваткиназная реакция. Последовательность реакций спиртового брожения идентична последовательности реакций гликолиза на всех этапах вплоть до образования пирувата, однако при спиртовом брожении пируват не восстанавливается до лактата, а декар-боксилируется с образованием ацеталь-дегида, который затем восстанавливается до этанола за счет NADH в реакции, катализируемой алкогольдегидрогена-зой. [26]
В функциональном отношении особо важное значение для вспышки гликолиза в мышце имеет активация фосфофруктокиназы продуктами реакции. Как видно из рис. 11 и 12, оба продукта - фруктозодифосфат и АДФ - активируют мышечную фосфофруктокиназу. Так как экспоненциальные вспышки гликолиза свойственны только мышце, не удивительно, что у других фосфофруктокиназ, например из печени, эритроцитов или кишечника, эта регуляторная особенность отсутствует или менее выражена. [27]
Оба этих метаболита ингибируют глюкозо-6 - фосфатде-гидрогеназу. В описанных опытах глюкозо-6 - фосфатдегидрогеназа выступает как регулируемый фермент пути Энтнера-Дудорова: функция этого фермента аналогична функции фосфофруктокиназы в фруктозо-бисфосфатном пути. [28]
Таким образом, эти реакции по своей природе преадап-тированы для регуляторной функции. Действительно, у большинства видов и в большинстве тканей три упомянутых фермента оказались главными пунктами регуляции гликолиза; из них фосфофруктокиназа обычно играет преобладающую роль в активации гликолитического процесса. [29]
Активность фосфофруктокиназы ингибируется также цитратом. Показано, что при диабете, голодании и некоторых других состояниях, когда интенсивно используются жиры как источник энергии, в клетках тканей содержание цитрата может возрастать в несколько раз. В этих условиях происходит резкое торможение активности фосфофруктокиназы цитратом. [30]
Страницы: 1 2 3 4