Фотосистема
Cтраница 2
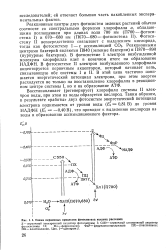 |
Схема первичных процессов фотосинтеза высших растений. [16] |
В фотосистеме I электрон возбужденной молекулы хлорофилла идет в конечном итоге на образование НАДФН. В фотосистеме II электрон возбужденного хлорофилла акцептируется первичным акцептором, который начинает цепь, связывающую обе системы I и II. В этой цепи частично понижается энергетический потенциал электрона, при этом энергия расходуется не только на восстановление хлорофилла в реакционном центре системы I, но и на образование АТФ. [17]
Как работают фотосистемы I и II при фотосинтезе. [18]
Следующий компонент фотосистемы 2 - цитохром 63 ( 6559) - представляет собой прочно связанный с ла-меллами хлоропластов хромопротеид. [19]
Искусственным вариантом фотосистемы 1 может быть восстановление экзогенной системы ферре-доксин - - НАДФ с помощью, например, дихлорфенол-индофенола восстановленного. [20]
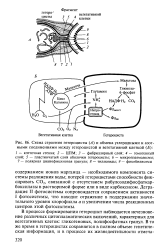 |
Схема строения гетероцисты ( А и обмена углеродными и азотными соединениями между гетероцистой и вегетативной клеткой ( Б. [21] |
Деградация II фотосистемы сопровождается сохранением активности I фотосистемы, что находит отражение в поддержании значительного уровня хлорофилла а и увеличении числа реакционных центров этой фотосистемы. [22]
Реакционный центр фотосистемы II содержит Хл а ( Р680), который служит первичным донором электронов во второй фотореакции. Получив энергию, поглощенную светособирающими пигментами фотосистемы II, этот хлорофилл переходит в возбужденное состояние. Этот электрон принимает молекула особого пластохинона ( Х320), который при этом восстанавливается до семи-хинона. Донором электронов для фотосистемы II служит вода. Разложение воды происходит при участии марганца. [23]
Рассмотрим компоненты фотосистем I и II зеленых растений и связывающих их звеньев. Последний является переносчиком с наибольшим из известных восстановительным потенциалом. Он представляет собой водорастворимый белок с М к 10000, содержащий линейно расположенные ионы железа, связанные друг с другом сульфидными группами, часть которых принадлежит цистеину. Именно эта часть белка является редокс-центром. Ферредоксины различных фото-синтезирующих объектов различаются по молекулярной массе, содержанию железа, значению редокс-потенциала. Все они обладают в окисленном состоянии красновато-желтой окраской - максимумы их полос лежат в областях 280, 330, 420 и 470 нм. [24]
Катион-радикал хлорофилла фотосистемы I восстанавливается, по-видимому, цитохромом f ( Цит. [25]
Получив электрон от фотосистемы I и ион Н от фотолиза воды, НАДФ превращается в НАДФ-Н, вступающий затем как восстановитель в процесс ассимиляции СО2 в синтезе глюкозы ( см. стр. [26]
 |
Схематическое изображение поверхности фотосистемы в тилако-идных мембранах ( цит. по А. Ленинджеру. [27] |
Все пигментные молекулы данной фотосистемы поглощают фотоны, но только одна молекула в каждом ансамбле обладает способностью превращать световую энергию в химическую. Она связана с особым белком - это и является фотохимическим центром. Все остальные пигментные молекулы - это светособирающие, или антенные, молекулы. [28]
Когда реакционный центр фотосистемы I, представляющий собой комплекс молекулы хлорофилла а с особым белком, переходит в возбужденное состояние под действием квантов света, полученных от антенных молекул, поглощение хлоропластов в области 700 нм снижается. [29]

Страницы: 1 2 3 4 5