Обратимый ингибитор
Cтраница 2
При таком механизме процесса ингибирующее влияние продуктов реакции может рассматриваться как частный случай действия конкурентных обратимых ингибиторов. Не исключено, однако, что продукт ферментативной реакции имеет возможность взаимодействовать, помимо того, с функциональными группами вне активного центра и, таким образом, влиять на активность фермента по принципу неконкурентного ингибитора. [16]
Это означает, что предположение о действии на группировки активного центра можно считать правильным, если действие обратимого ингибитора конкурентно в отношении субстрата или если реакция с необратимым ингибитором в присутствии субстрата замедляется. [17]
По-видимому, исторически сложилось так, что применение величины / во для характеристики необратимых ингибиторов было некритически заимствовано из опыта исследования обратимых ингибиторов. Возможно, что при этом важную роль играла недостаточная изученность механизма действия необратимых ингибиторов и стремление провести аналогию с действием обратимых ингибиторов. Позднее накопился большой экспериментальный материал по характеристике необратимых ингибиторов с использованием / 8о в качестве меры их антиферментной активности. Любопытно, что в некоторых случаях эта мера позволяла проводить сопоставление между химическим строением и антиферментным действием в рядах родственных по строению ингибиторов. По-видимому, это объясняется тем, что в подобных случаях время взаимодействия ингибитора с ферментом в среднем у разных авторов не слишком различалось и реакция не доходила до конца. Однако при более тщательном исследовании возникли, как и следовало ожидать, явные противоречия, объясняющиеся неадэкватностью параметра / 6о для необратимых ингибиторов. [18]
В числе всех этих так называемых функциональных противоядий мы рассмотрели вещества, препятствующие действию яда на рецепторные структуры или блокирующие это действие ( атрошшоподобные соединения, адрено-блокаторы, обратимые ингибиторы ферментов); реактивирующие биоструктуры с последующей окончательной ЕЛИ временной детоксикацией яда ( оксимы, унитиол, цистамин); конкурирующие с ядом за связь с рецепторами ( кислород, структурные аналоги токсичных лекарств); замещающие биоструктуры поврежденные или утраченные под воздействием яда ( ниридоксин, цитохром Ct: хлорид кальция); активаторы и ингибиторы ферментовf ыетаболизирующих яды ( препарат SKF - 525A, антиокси-данты), и ряд других. Некоторые из них являются антидотами многостороннего действия и, значит, могут использоваться при различных отравлениях или же воздействовать на один и тот же токсический процесс разными путями. Например, глюкоза и метиленовый синий - анти - Еианнды и в то же время восстановители метгемоглобина. Кроме того, глюкоза может быть заместительным антидотом при инсулиновой интоксикации, а метиленовый синий влиять на процессы тканевого окисления, нарушенные ядами. Здесь уместно отметить, что в последнее время все большее значение в качестве противоядий многостороннего действия приобретают антиоксиданты. [19]
Об одинаковом влиянии концентрации тетраметиламмония на константы скорости реакции всех трех необратимых ингибиторов свидетельствуют данные табл. 32, показывающие снижение величины константы ( в %) при увеличении концентрации ионов обратимого ингибитора. Подобные результаты были получены для тетраэтил-аммоний-иона; это позволило предположить, что все три ФОС в присутствии ионов тетраалкиламмония реагируют лишь с полностью свободными активными центрами и не реагируют с нуклео-фильными группировками ферментов, если анионная группа занята ионом тетраалкиламмония. Иными словами, реакционная способность нуклеофильной группировки к ФОС полностью утрачивается, если анионная группировка занята ионом тетраалкиламмония. [20]

В опытах с армином ( синтезирован А. И. Разумовым в КХТИ) было показано, что характерные реакции тонических и нетонических мышц на воздействие фосфорорганическими веществами принципиально не отличаются от их реакций на обратимые ингибиторы типа эзерина. [22]
Несомненно, что анионная группировка в активном центре аце-тилхолинэстеразы играет более важную роль, чем в холинэстеразе при каталитическом действии фермента, и, как будет показано дальше, это проявляется не только в случае обратимых ингибиторов. [23]
Таким образом, согласно этому уравнению, если верно предположение, что в присутствии обратимого ингибитора необратимый ингибитор не реагирует с какими-либо группировками активного центра, то между обратной величиной бимолекулярной константы скорости реакции фермента с необратимым ингибитором и концентрацией обратимого ингибитора должна быть линейная зависимость. [24]
Обратимое ингибирование характеризуется равновесием между ферментом и ингибитором Константа равновесия ( Kt) служит мерой их сродства. Обратимые ингибиторы, действие которых приводит к увеличению эффективной Кт, называют конкурентными. Неконкурентные ингибиторы не влияют на величину Кт, но уменьшают максимальную скорость. [25]
Обратимое ингибирование поддается количественному изучению на основе кинетики Михаэлиса-Ментен. Обратимые ингибиторы делят на конкурентные и неконкурентные. [26]
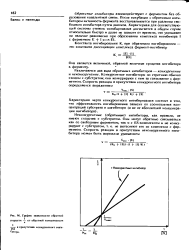 |
График зависимости обратной. [27] |
Обратимые ингибиторы взаимодействуют с ферментом без образования коаалентиой связи. После инкубации с обратимым ингибитором активность фермента восстанавливается при удалении свободного ингибитора путем диализа. [28]
Обладает антюсолинэстеразной активностью, являясь прямым обратимым ингибитором фермента. [29]
В обзоре [84] рассмотрены типы биосенсоров, амперометрический отклик которых обусловлен восстановлением тиолов, образующихся при гидролизе тиохолина. Они позволяют определять в основном обратимые ингибиторы. [30]
Страницы: 1 2 3 4