Обратимый ингибитор
Cтраница 3
В обзоре 84 ] рассмотрены типы биосснсоров, амперометрический отклик которых обусловлен восстановлением тиолов, образующихся при гидролизе тиохолина. Они позволяют определять в основном обратимые ингибиторы. [31]
Так, галантамин ( см. Амариллисовые алкалоиды) и др. обратимые ингибиторы холин-эстераз защищают эти ферменты от взаимод. [32]
Активация проингибитора и его превращение в ингибитор в отсутствие гемина проходит две фазы. Сначала, в течение минут, проингиби-тор без гемина превращается в так называемый обратимый ингибитор; добавление к нему гемина снова переводит его в состояние проингибитора. Природа этого превращения, обязательно требующего физиологической температуры, не ясна; так как общие физические параметры этого белка в состоянии проингибитора и обратимого ингибитора одинаковы, предполагают какую-то внутримолекулярную перестройку, приводящую к появлению ингибиторной активности. При дальнейшей инкубации, в течение часов, обратимый ингибитор превращается в необратимый ингибитор. Наоборот, SH-соединения типа восстановленного глютатиона, р-меркаптоэтанола и дитиотреита препятствуют превращению обратимого ингибитора в необратимый ингибитор. При превращении происходит также автофосфорилирование ингибитора за счет АТФ. [33]
По-видимому, исторически сложилось так, что применение величины / во для характеристики необратимых ингибиторов было некритически заимствовано из опыта исследования обратимых ингибиторов. Возможно, что при этом важную роль играла недостаточная изученность механизма действия необратимых ингибиторов и стремление провести аналогию с действием обратимых ингибиторов. Позднее накопился большой экспериментальный материал по характеристике необратимых ингибиторов с использованием / 8о в качестве меры их антиферментной активности. Любопытно, что в некоторых случаях эта мера позволяла проводить сопоставление между химическим строением и антиферментным действием в рядах родственных по строению ингибиторов. По-видимому, это объясняется тем, что в подобных случаях время взаимодействия ингибитора с ферментом в среднем у разных авторов не слишком различалось и реакция не доходила до конца. Однако при более тщательном исследовании возникли, как и следовало ожидать, явные противоречия, объясняющиеся неадэкватностью параметра / 6о для необратимых ингибиторов. [34]
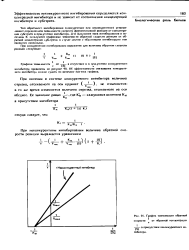 |
График зависимости обратной. [35] |
Тип обратимого ингибирования ( конкурентное или неконкурентное) устанавливают определением зависимости скорости ферментативной реакции от концентрации субстрата в присутствии ингибитора. Дчя выяснения типа ингибирования и величины К пользуются графиками зависимости обратной скорости реакции от обратной концентрации субстрата; в стучае систем с обратимыми ингибиторами они являются линейными. [36]
Определение обратимых ингибиторов холинэстераз менее распространено. Тем не менее в настоящее время разработано множество тестов на основе холинэстераз для определения ионов тяжелых металлов и хло-рорганических пестицидов [89,90], являющихся обратимыми ингибиторами. Интерес представляет способ оценки общей токсичности водопроводной воды, основанный на суммарном эффекте снижения ферментативной активности холинэстеразы при введении в систему субстрат-фермент анализируемой пробы. [37]
Определение обратимых ингибиторов холинэстераз менее распространено. Тем не менее в настоящее время разработано множесгво тестов на основе холинэстераз для определения ионов тяжелых металлов и хло-рорганических пестицидов 89 90), являющихся обратимыми ингибиторами. Интерес представляет способ оценки общей токсичности водопроводной воды, основанный на суммарном эффекте снижения ферментативной активности холинэстеразы при введении в систему субстрат-фермент анализируемой пробы. [38]
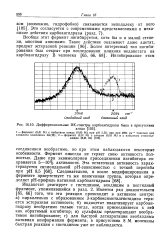 |
Дифференциальные ИК-спектры карбоангидразы быка в присутствии. [39] |
Иодацетат реагирует с гистидином, входящим в пептидный фрагмент, упоминавшийся в разд. Имеется ряд доказательств [65, 66] того, что эта реакция происходит в активном центре: а) параллельно с образованием 3-карбоксиметилгистидина теряется эстеразная активность; б) иодацетат ивактивирует карбоангидразу необратимо только тогда, когда присоединяется к ней как обратимый ингибитор; в) сульфиды предотвращают необратимое ингибирование; г) для быстрой дегидратации необходима нативная конформация фермента; в присутствии 8 М мочевины быстрая реакция с гистидином уже не наблюдается. [40]
Ингибитор в активированном состоянии оказался обладающим ц АМФ-независимой протеинкиназнои активностью, специфически фос-форилирующей а-субъединицу eIF - 2 за счет АТФ. Именно эта активность и является ответственной за ингибирование ( репрессию) инициации трансляции в клетках, лизатах, экстрактах и бесклеточных системах ретикулоцитов в отсутствие гемина. Киназная активность обратимого ингибитора подавляется гемином. Ингибитор, как уже отмечалось, способен также к автофосфорилированию, которое, возможно, играет какую-то роль в регуляции его активности. [41]
При этом было установлено, что величина / so лишь в случае чисто неконкурентных ингибиторов совпадает с / G. Уже для конкурентных обратимых ингибиторов при переходе от 5о к Kt необходим учет величин константы Михаэлиса и концентрации субстрата, поскольку в системе устанавливается равновесие между Е, S и I ( стр. Что касается необратимых ингибиторов, то в этом случае использование / 8о для оценки их реакционноспособ-ности без учета времени реакции не имеет никаких оснований. [42]
Ингибиторы ферментативных каталитических реакций часта подразделяют на обратимо и необратимо действующие. Эта классификация основана на легкости отделения ингибитора от фермента при помощи физического метода типа диализа. Так, например, эзерин описан как обратимый ингибитор, в то время как фосфорсодержащие соединения подобно диизопропилфосфофториду ( DPF) принадлежат к необратимым ингибиторам холинэстеразы. Однако, поскольку рассматриваются механизмы ингибирования, эта классификация только вносит неопределенность, так как из нее вытекает, что обратимые и необратимые ингибиторы действуют различным образом; в действительности оба типа ингибиторов действуют, соединяясь с ферментом с образованием неактивных комплексов, обладающих весьма различными константами диссоциации. Необратимые ингибиторы образуют комплексы с очень малыми константами диссоциации и поэтому удаляются из комплекса путем диализа только очень медленно, тогда как обратимые ингибиторы образуют комплексы с высокими константами диссоциации и поэтому на всех стадиях удаления присутствует избыток несвязанного ингибитора, поддающегося диализу. Однако более полезным оказывается подразделение ингибиторов на конкурентные и неконкурентные ( хотя многие ингибиторы обнаруживают смешанное поведение), так как эти ингибиторы действуют различными путями и кинетические уравнения для них разные. [43]
Подход к измерению константы продукта может быть двояким. Продукт реакции вводится заранее в систему фермент - субстрат, после чего измеряется зависимость начальной стационарной скорости реакции в ряду концентраций субстрата и продукта. Этот способ фактически ничем не отличается от способа исследования обычных обратимых ингибиторов. [44]
До открытия антихолинэстеразных свойств органических соединений фосфора существовало лишь сравнительно немного фармацевтических препаратов, обладающих хо-линэргическим действием. К их числу относились такие алкалоиды, как никотин, производные ацетилхолина, простигмин и другие родственные ему карбаматы. Все эти препараты представляли собой либо непосредственные стимуляторы холинорецепторов, либо обратимые ингибиторы холинэстеразы, активность которых, следовательно, являлась временной в противоположность необратимому по существу действию органических соединений фосфора. [45]
Страницы: 1 2 3 4