Углеводная цепь
Cтраница 3
Гликозидные связи устойчивы к действию щелочей, о если олиго - или полисахарид содержит восстанавливающий сахар, происходит деструкция углеводной цепи, начиная с этого восстанавливающего остатка. [31]
Усложнение углеводной цепи О-гликозилпротеинов происходит в результате присоединения к кору остатков М - ацетиллактозамина Са1р1 - 4С1сМАср1 - ; на конце углеводных цепей находятся обычно остатки фукозы или М - ацетилнейраминовой к-ты. [32]
 |
НаиГолее распространенные символы важнейших моносахаридов. [33] |
Особенно широко эта система записи используется в работах, связанных с исследованиями полисахаридов, а также в других областях, требую-цих записи достаточно длинных углеводных цепей. [34]
Многие полисахариды, в том числе крахмал и целлюлоза, существуют в природе в кристаллическом и полукристаллическом состояниях, и, следовательно, направление реакций замещения и модификации этих соединений определяется формой их молекул и способом укладки углеводных цепей. [35]
Второй гликопротеин куриного белка - овомукоид имеет молекулярный вес примерно 28 000 и содержит около 20 % углеводов, в числе которых находится манноза, галактоза, глюкозамин и небольшие количества N-ацетилнейраминовой кислоты. Строение углеводных цепей - овомукоида еще не установлено, однако в результате протеолиза59 и частичного кислотного гидролиза выделено несколько гликопептидов. [36]
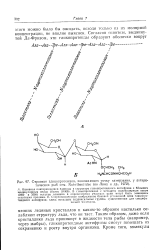 |
Строение глпкопротеидов, понижающих точку замерзания, у антарктических рыб сем. Nototheniidae ( по Лину и др., 1972. [37] |
В гликопротеидах с меньшим молекулярным весом ( 2600 и 3500) остатки аланина в определенных участках цепи могут быть замещены остатками пролнна. Стереохимичсское изображение боковой углеводной цепи гликопротеидного антифриза; здесь показаны гндроксильные группы, существенные для специфической молекулы. [38]
Боковые углеводные цепи гликопро-теинов ковалентно присоединяются к полипептиду во время или после синтеза последнего. В одних гликопротеинах боковая углеводная цепь прикрепляется с помощью ферментов к остаткам аспа-рагиновой кислоты, в других - серина или треонина. Многие белки, которые работают вне клетки, а также смазочные протеогликаны, покрывающие слизистые оболочки, содержат боковые олигосаха-ридные цепи. [39]
Овальбумин является наиболее доступным54 гликопротеином, послужившим первым объектом серьезного химического изучения гликопротеинов. Овальбумин содержит одну углеводную цепь и относится поэтому к гликопротеинам типа I ( см. стр. [40]
Молекулы фосфолипидов, гликолипидов и гликопротеинов также могут транспортироваться. Если радиоактивно меченная фукоза, компонент углеводных цепей многих гликопротеинов, вводится в тело клетки, то она включается в белок в цитоплазме, а не в нервном окончании. В то же время N-ацетилнейраминовая кислота ( NANA) ведет себя по-другому: она, видимо, встраивается в гликопротеин или гликолипид только в нервном окончании. [41]
Определение мест привязки олигосахаридных цепей в белках проводится путем подбора таких условий расщепления полипептидной цепи, которые позволили бы получить фрагменты, несущие лишь по одной углеводной цепочке. После определения последовательности аминокислот и положения углеводной цепи в глико-пептиде локализуют его положение в исходной молекуле. [42]
Направленное гликозилирование белков, например, ре комби-нантных, полученных в бактериях с целью создания точных копий природных белков, пока неосуществимо ни химическим, ни ферментативным способом. Причина заключается в сложности процесса биосинтеза углеводных цепей: его осуществляет сложный комплекс ферментов - гликозилтрансфераз, каждый из которых присоединяет определенный углеводный остаток. Кроме того, присоединение углеводов начинается уже в процессе биосинтеза полипептидной цепи гликопротеина, поэтому место будущей углеводной цепи оказывается помеченным еще до того, как произойдет формирование третичной структуры белка. В сформировавшейся глобуле места потенциального гликозилирования могут быть недоступными для действия соответствующих ферментов. [43]
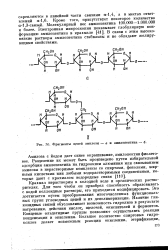 |
Фрагменты цепей амилозы - а и амилопектина - б. [44] |
Крахмал нерастворим в холодной воде и органических растворителях. Это достигается путем преобразования многочисленных функциональных групп углеводных цепей и их деполимеризации. Наличие гли-козидных связей обусловливает возможность гидролиза в результате нагревания, действия кислотл щелочей, окислителей и ферментов. Концевые альдегидные группы позволяют осуществлять реакции конденсации и окисления. [45]
Страницы: 1 2 3 4