Полинуклеотидная цепь
Cтраница 2

Синтез одной полинуклеотидной цепи таким способом объяснить нетрудно: ДНК-полимераза способна непрерывно связывать нуклеотиды по направлению от 5 к З - концу. Но так как двойная спираль ДНК антипараллельна, синтез второй, противолежащей цепи должен происходить в обратном направлении. Эти соображения и экспериментальные данные привели к представлению, которое иллюстрируют схемы на рис. 2.16. Вероятно, сначала образуются только короткие ( длиной около 1000 нуклеотидов) отрезки-так называемые фрагменты Оказаки. [17]
Электронные переходы полипептидных и полинуклеотидных цепей и, тем самым, белков и нуклеиновых кислот расположены в ультрафиолетовой области спектра. Полосы поглощения пептидной связи - СО-NH - лежат в области 185 - 240 им, здесь же и в более коротковолновой области расположены полосы алифатических боковых цепей аминокислотных остатков. Ароматические остатки Три, Фен, Тир имеют полосы поглощения в области 280 нм. Азотистые основания в нуклеиновых кислотах поглощают свет в области 260 нм. Таким образом, белки и нуклеиновые кислоты бесцветны, они не поглощают видимый свет. [18]
 |
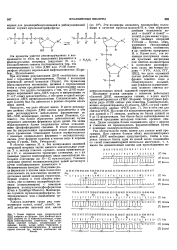
Двойная спираль ДНК. [19] |
Сахарофосфатные остовы двух полинуклеотидных цепей, расположенные снаружи, связаны между собой водородными связями между отходящими от них вовнутрь азотистыми основаниями. Плоскости оснований перпендикулярны оси спирали и отстоят друг от друга на 0 34 нм. [20]
Решение проблемы синтеза полинуклеотидных цепей достигнуто путем комбинирования химических и биохимических методов. [22]
Концевая фосфатная группа полинуклеотидной цепи может быть превращена в фосфоанилид взаимодействием с 14С - анилином и дициклогексилкарбодиимидом197; более общий метод состоит в превращении концевого остатка нуклеотида в двузамещенный пи-рофосфат 198 при реакции с 14С - метилфосфоморфолидом. [23]
Конечным нуклеотидом в полинуклеотидной цепи транспортной РНК, реагирующим с аминоациладенилатом, всегда является адениновый нуклеотид. [24]
Теоретические расчеты копформаций полинуклеотидных цепей и двойных спиралей по просты, так как эти конформации зависят от многих углов. Копформациопно лабильна и группа рибозы или дезоксирибозы. Соответственно проводятся расчеты с жестким и мягким сахаром. [25]
Конденсация рибонуклеотидов в полинуклеотидную цепь ( точно таким же образом, как в случае ДНК) приводит к РНК. Различные РНК имеют молекулярную массу от 20000 до 200000 и более. В качестве оснований они содержат остатки аденина, гуанина, цитозина, урацила я в незначительных количествах остатки гипоксантина. Тимин в РНК в качестве основного компонента не встречается. Большинство типов РНК имеет однотяжевую структуру. В зависимости от нахождения в клетке и функции различают рибосомные РНК, ( pPHK) f транспортные РНК ( тРНК), информационные РНК ( uPHK) f ядерные РНК. [26]
Влияние рН на конформации полинуклеотидных цепей в растворе обусловлено тем обстоятельством, что водородные связи, стабилизующие спиральную структуру, образуются в этих молекулах между группами, способными к ионизации, и поэтому ионизация хотя бы одной из групп, участвующих в образовании водородной связи, означает одновременно разрыв последней, что ведет к изменению конформации молекулы. Пуриновые и пиримидиновые основания, входящие в ДНК и синтетические полинуклеотиды, образуют водородные связи между аминогруппой и атомом азота, включенным в цикл, с одной стороны, и группой - NH-СО - с другой. Поскольку аминогруппа присоединяет протон, а группа - NH-СО - отдает его, то первая заряжена при рН jo / NHz а вторая при рН рК и-со - Таким образом, в диапазоне рКт2 С рН рКхн - - со пуриновые и пиримидиновые основания не заряжены, и здесь возможно существование спиральной конформации молекул. [27]
Вопрос о влиянии длины полинуклеотидной цепи на скорость гидролиза в настоящее время еще не ясен. [28]
Все изложенное относилось к полинуклеотидным цепям. Мы видим, что сколько-нибудь длинные цепи ( по оценке Эйгена с v 100) сами по себе не эволюционируют. [29]
Давайте предположим, что имеется полинуклеотидная цепь, заканчивающаяся последовательностью фXYфZ, и что наряду с другими нуклеотидами в нее включается уридин-5 - монофосфат. Введенный в молекулу УМФ может быть расположен либо на конце молекулы - фХфУф7фУ, либо где-то внутри нее, например - фХфУфУф2 ( фиг. Щелочной гидролиз позволяет получить затем смесь нуклеозид-3 ( 2) - монофосфатов и один-единственный нуклеозид, образованный из концевого нуклеотида. Если УМФ был помечен С14 по основанию и присоединился к концу полирибонуклеотидной цепи, то после щелочного гидролиза меченым окажется единственный нуклеозид - уридин. Если же УМФ включается в середину молекулы, то при гидролизе радиоактивность будет найдена в уридин-3 ( 2) - монофосфате. [30]
Страницы: 1 2 3 4