Гель-электрофорез
Cтраница 2
Амплифицированный фрагмент обрабатывают Cvnl, продукты рестрикции разделяют с помощью гель-электрофореза и окрашивают их бромистым этидием. [16]
Полисахарид, нейтральный компонент агара; используется как носитель для гель-хроматографии и гель-электрофореза биополимеров. [17]
Одним из наиболее изученных 4 ферментов, множественность форм которого детально изучена методом гель-электрофореза, является ЛДГ, катализирующая обратимое превращение пировиноградной кислоты в молочную. Пять изоферментов ЛДГ образуются из 4 субъединиц примерно одинакового размера, но двух разных типов. [18]
Затем все четыре образца параллельно разгоняются в одном и том же аппарате для гель-электрофореза. [19]
Число витков суперспирали определяется различными способами - косвенно по связыванию лигандов и непосредственно методом гель-электрофореза. Этот метод очень чувствителен, он позволяет определить молекулы, у которых т разнятся всего лишь на единицу. [20]
Недавно было показано, что нуклеопротеиды [147] и другие белки [148], разделенные с помощью гель-электрофореза, можно перенести на нитроцеллюлозные или диазобензилоксиметшщел-люлозные фильтры. [21]
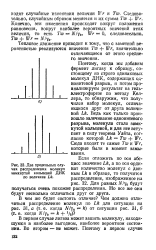 |
Два предельных случая распределения молекул замкнутой кольцевой ДНК по величине Lk. [22] |
Поэтому, когда мы добавим фермент лигазу к образцу, состоящему из строго одинаковых молекул ДНК, содержащих од-нонитевой разрыв, и потом проанализируем результат на гель-электрофорезе по методу Келлера, то обнаружим, что получился набор молекул, отличающихся друг от друга величиной Lk. Ведь как только произошло залечивание однонитевого разрыва, молекула стала замкнутой кольцевой, и для нее вступает в силу теорема Уайта, согласно которой Lk Tw - f - Wr. Сюда входят те самые Tw и Wr, которые были в момент замыкания. [23]
 |
Карта линейного расположения пистонов на ДНК. [24] |
Один из этих подходов состоит в локализации ковалентных сшивок между нуклеосомными гистонами и ДНК - Принцип локализации заключается в том, что сшитые комплексы белков с ДНК разделяют в двумерных гель-электрофорезах, причем после электрофореза первого направления в геле расщепляют белковый или нуклеиновый компонент комплексов и разделяют во втором направлении только ДНК или только белки соответственно. Гистоны НЗ, Н4 располагаются в центре нуклеосомной ДНК, в то время как гистоны Н2А, Н2В локализованы на периферии. Гистон НЗ взаимодействует с центральным и концевым участками нуклеосомной ДНК. Хотя эти участки на развернутой ДНК расположены далеко друг от друга, они сближаются на свернутой в нуклеосому ДНК и, видимо, с ними взаимодействует одна и та же молекула гистона НЗ. На этой же карте видно, что не вся ДНК сплошь покрыта гистонами, а есть свободные от взаимодействия сегменты, например первые 20 нуклеотидов от 5 -концов обеих цепей нуклеосомной ДНК и участки, расположенные на расстоянии около 120 нуклеотидов от 5 -концов. [25]
 |
Карта линейного расположения гистонов на ДНК. [26] |
Один из этих подходов состоит в локализации ковалентных сшивок между нуклеосомными гистонами и ДНК - Принцип локализации заключается в том, что сшитые комплексы белков с ДНК разделяют в двумерных гель-электрофорезах, причем после электрофореза первого направления в геле расщепляют белковый или нуклеиновый компонент комплексов и разделяют во втором направлении только ДНК или только белки соответственно. Гистоны НЗ, Н4 располагаются в центре нуклеосомной ДНК, в то время как гистоны Н2А, Н2В локализованы на периферии. Гистон НЗ взаимодействует с центральным и концевым участками нуклеосомной ДНК. Хотя эти участки на развернутой ДНК расположены далеко друг от друга, они сближаются на свернутой в нуклеосому ДНК и, видимо, с ними взаимодействует одна и та же молекула гистона НЗ. На этой же карте видно, что не вся ДНК сплошь покрыта гистонами, а есть свободные от взаимодействия сегменты, например первые 20 нуклео-тидов от 5 -концов обеих цепей нуклеосомной ДНК и участки, расположенные на расстоянии около 120 нуклеотидов от 5 -концов. [27]
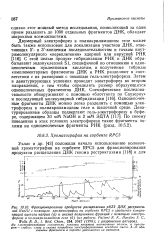
С этой целью продукт гибридизации РНК и ДНК обрабатывают эндонуклеазой S1, специфичной по отношению к одно-цепочечным нуклеиновым кислотам, и полученные гибриды комплементарных фрагментов ДНК и РНК разделяют в соответствии с их размерами с помощью гель-электрофореза. Затем в денатурирующих условиях проводят электрофорез во втором направлении, с тем чтобы определить размер образующихся одноцепочечных фрагментов ДНК. Одноцепочеч-ные фрагменты ДНК можно легко разделить с помощью электрофореза в полиакриламидном геле в щелочной среде. По своему поведению в этих условиях электрофореза такие фрагменты похожи на одноцепочечные фрагменты РНК ( разд. [29]
Метод гель-электрофореза позволяет выделить каждый фрагмент в изолированном виде. [30]
Страницы: 1 2 3 4