Гидролиз - гликозидная связь
Cтраница 2
 |
Принцип плюс и минуса-метода определения последовательности. [16] |
Эта ситуация легко меняется на обратную, когда достигается сильное расщепление по аденину и слабое по гуанину, при использовании для гидролиза гликозидной связи слабокислых условий, в которых подавляется гидролиз 7-метилгуанозина. [17]
До последнего времени не решена альтернатива о том, проходят ли реакции, катализируемые лизоцимом, через образование кова-лентного гликозилфермента ( двухстадийный гидролиз гликозид-ной связи) или осуществляется одностадийный механизм гидролиза гликозидной связи. [18]
Применение этих методов к полисахаридам с высоким молекулярным весом требует соблюдения условий, в которых образование формальдегида и углекислого газа протекало бы довольно быстро, но не происходило бы ни гидролиза гликозидных связей, ни сколько-нибудь существенного случайного окисления макромолекулы. Было найдено, что в этих условиях экви-молярные количества мальтодекстринов переокисляются с совершенно одинаковой скоростью. [19]
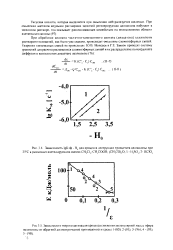 |
Зависимость 1 § К. эф - Н ( 1 для процесса деструкции триацетата целлюлозы при 25 С в различных ацетилирующих смесях СН С1. СН. СООН. ( СН. СО О. I - Н 8О. 2 - НС1О4. [20] |
При обработке волокна частично-замещенного ацетата ( диацетата) целлюлозы растворами оснований, как было уже сказано, происходит омыление сложноэфирных связей. Гидролиз гликозидных связей не происходит. [21]
Место связи фруктозного и глюкозного остатков друг с другом доказывается посредством метилирования всех свободных гидроксилов сахарозы. Гидролиз гликозидной связи продукта метилирования дает два частично метилированных моносахарида. [22]
Место связи фруктозного и глюкозного остатков друг с другом доказывается посредством метилирования всех свободных гидроксилов сахарозы. Гидролиз гликозидной связи продукта метилирования дает два частично метилированных моносахарида. Положения свободных гидроксильных групп в этих соединениях соответствует местам соединения двух моносахаридных остатков в дисахариде. [23]
Известные методы расщепления N-гликозидных связей в нуклеозидах, нуклеотидах и нуклеиновых кислотах можно условно подразделить на две группы: 1) прямые методы расщепления N-гликозидных связей в обычных нуклеозидных звеньях под действием кислот или, реже, оснований; 2) косвенные методы, в которых лабилизация N-гликозидных связей достигается предварительной модификацией гетероциклического основания или сахарного остатка. Реакции гидролиза гликозидных связей после модификации оснований, не приводящей к нарушению их ароматического характера, мы относим к прямым методам. [24]
Этот механизм - последовательность стадий, обратная процессу образования ацеталей из альдегидов и спиртов. При гидролизе гликозидной связи как ацеталя алкоксильная группа является плохо уходящей группой, что и вызывает необходимость ее предварительного протониро-вания с переводом в сопряженную кислоту. [25]
При обработке волокна ДАЦ растворами оснований в результате гидролиза слож ноэфирных связей образуется слой регенерированной целлюлозы. При этом гидролиза гликозидных связей не происходит. [26]
Выбор кислоты и ее концентрации зависит от вида гидролизуе-мого образца и цели гидролиза. Кроме показателя рКа на скорость гидролиза гликозидных связей влияют гидролитическая активность ( выражаемая значением рН), коэффициент активации иона гидроксония ( выражаемый функцией кислотности Гаммета), а также температура и давление. [27]
Положение фосфатной группы у этих двух изомеров было первоначально приписано на основании данных физических измерений и их гидролиза, в результате которого образуется D-рибозо - 2 -фосфат и З - фосфат. Тем не менее, однозначный синтез этих соединений оказался необходимым, поскольку выяснилось, что внутримолекулярное взаимопревращение этих двух изомеров, катализируемое кислотой, происходит со скоростью, сравнимой со скоростью гидролиза гликозидной связи. Фосфат ( 46) был идентичен а-изомеру [57], который элюировался первым с колонки Кона. Аналогичный синтез продемонстрировал, что уридиловая кислота а является 2 -фосфатом. [28]
Принципиальным преимуществом ферментативного гидролиза перед кислотным является его специфичность, позволяющая не только устанав - ливать строение полисахарида по сохранившимся фрагментам, но - и по -, дучать определенные сведения о гликозидных связях, разорвавшихся при; гидролизе. Специфичность разных ферментов может различаться очень сильно. Некоторые из них катализируют гидролиз гликозидных связей определенной конфигурации нескольких родственных моносахаридов, другие специфичны как к кон - фигурации гликозидного центра, так и к остатку моносахарида, а третьи: требуют определенного расположения моносахаридов, удаленных от расщепляемой гликозидной связи. Так, например, амилазы обладают прак - гически абсолютной специфичностью, вызывая расщепление только 2 - 1 - - 4-связей D-глюкопираноз в крахмалоподобных полисахаридах107, и чрезвычайно чувствительны даже к незначительным изменениям структуры субстрата. [29]
Совершенно иначе эта реакция протекает в присутствии катализаторов, направляющих ее в сторону образования моносахаридов. Катализаторами являются кислоты, кислые соли и другие соединения, которые в водном растворе диссоциируют с образованием ионов водорода. Последний собственно и является катализатором гидролиза гликозидных связей. Каталитическая активность веществ, образующих в водном растворе ионы водорода, зависит от их силы или способности диссоциировать. Чем полнее вещество диссоциирует в воде, тем более сильным катализатором оно является. Так, кислоты по их каталитической активности образуют ряд, соответствующий их силе диссоциации. В этом ряду на первом месте стоят минеральные кислоты, на 100 % диссоциирующие в разбавленных водных растворах, например соляная кислота. Каталитическая активность серной кислоты почти в 2 раза меньше, так как ее второй водород имеет значительно меньшую константу диссоциации. [30]
Страницы: 1 2 3