Гиппокамп
Cтраница 1
Гиппокамп, находящийся под двумя главными мозговыми полушариями, видимо, занимается обработкой и распределением поступающей информации, которая хранится затем в коре мозга и мозжечке. [1]
Таким образом, гиппокамп не является хранилищем информации, а ответствен за консолидацию памяти или трансформацию кратковременной памяти в долговременную. [2]
Еще один класс нейронов гиппокампа заслуживает особого внимания. При действии раздражителя эти нейроны сначала отвечают с определенным запаздыванием. [3]
Не только неокортекс, но и гиппокамп играет, видимо, особую роль. [4]
Вероятно, в случае с был поврежден гиппокамп, поскольку память на прошлые события осталась ненарушенной, но образовать новые воспоминания было очень трудно. Исследования мозга указывают на то, что два типа памяти - процедурная и декларативная - связаны с этими главными структурами. Процедурная память хранит моторные навыки, такие как письмо от руки или на машинке или ( возможно) способность ездить на велосипеде; она располагается в основном в мозжечке. Декларативная память содержит сведения и знания о мире, такие как имя любимой тетушки, местонахождение ближайшей пиццерии, значения слов и целая уйма другой информации, и все это хранится в коре мозга. [5]
Следует подчеркнуть, что это угасание реакций пирамидных нейронов гиппокампа селективно в отношении параметров стимула. Важным условием привыкания реакций пирамидных нейронов на приход сигналов от коры является потенциация синапсов нейронов зубчатой фасции, также приходящих на пирамидные нейроны гиппокампа. Таким образом, сигналы корковых детекторов поступают параллельно, прямо на пирамидные нейроны и через нейроны зубчатой фасции. При этом нейроны зубчатой фасции селективно блокируют локальные участки дендритов пирамидных нейронов, куда приходят корковые сигналы. Соседние синапсы корковых детекторов генерируют в дендритах пирамидных нейронов возбуждающие постсинаптические потенциалы ( ВПСП), которые развиваются без шунтирующего взаимодействия между участками мембраны, где они возникают. Гаммааминома-сляная кислота, ионофоретически локально аплицируемая на дендрит пирамидного нейрона, вызывает деполяризацию, которая связана с локальным повышением проводимости на данном участке мембраны дендрита. В результате соседние ВПСП подвергаются депрессии. ВПСП, вызванные на удаленных участках дендрита под влиянием вызванной гаммааминомасляной кислотой деполяризации, оказываются более эффективными в отношении генерации потенциалов действия. Этот механизм предположительно функционирует при депрессии корковых входов нейронами зубчатой фасции. [6]
Нарушения интрацен-тральных отношений между мезэнцефалической ретикулярной, формацией и гиппокампом, гиппокампом и корой становятся более выраженными, нарушается характер реципрокных отношений между этими структурами, а характер интрацентральных отношений между мезэнцефалической ретикулярной формацией и корой не изменен. [7]
Опыты поставлены на бодрствующих кроликах с электродами, вживленными в гиппокамп, ретикулярную формацию среднего мозга и структуры обонятельного анализатора - обонятельные луковицы, пириформную кору, кортикомедиальное и латеральное ядра амигдалы. Установлено, что в случае применения функциональной нагрузки в виде ритмического света формальдегид в концентрации 0 65 мг / м3 ухудшает реакцию перестройки ритма в кортико-медиальном ядре амигдалы. Концентрация 0 035 мг / м3 действия на электропотенциалы кроликов не оказывает. [8]
Если гипотеза о конвергенции множества детекторов на каждом интегративном нейроне гиппокампа правильна, то следует ожидать, что характеристика селективного угасания гиппокампального интегративного нейрона должна воспроизводить селективность реакции возбуждаемых стимулом детекторов. [9]
Нарушения интрацен-тральных отношений между мезэнцефалической ретикулярной, формацией и гиппокампом, гиппокампом и корой становятся более выраженными, нарушается характер реципрокных отношений между этими структурами, а характер интрацентральных отношений между мезэнцефалической ретикулярной формацией и корой не изменен. [10]
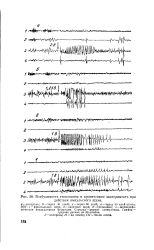 |
Возбудимость гиппокампа в хроническом эксперименте при. [11] |
ЭЭГ: 1 - фронтальная кора; 2-височная кора; 3 - гиппокамп; 4 - меЗэнцефа - лическая ретикулярная формация. [12]
С этой точки зрения нервная модель стимула строится параллельно на множестве интегративных нейронов гиппокампа за счет трансформации отдельных синаптических контактов, образованных детекторами на интегративном нейроне в виде матрицы. Внешний стимул отображается на этой матрице посредством детекторов в виде конфигурации трансформированных локусов. Смена раздражителя означает перемещение действующих локусов на мембране нейрона в соответствии с тем, на какие детекторы этот стимул воздействует. [13]
Из табл. I видно, что содержание 3Н - лейцина в нервных клетках гиппокампа ( имп / мин надосадочной жидкости на мкг белка в образце) у обучавшихся животных ниже, чем у контрольных. Это можно объяснить потреблением 3Н - лейцина во время обучения, что приводит к тому, что количество доступного 3Н - лейцина во время обучения ниже того, которое должно быть. [14]
Посттетаническая потенциация может длиться от нескольких минут до многих часов ( в синапсах гиппокампа) и играть важную роль в пластических изменениях функции синапсов. [15]
Страницы: 1 2 3 4