Гиппокамп
Cтраница 4
Степень выраженности судорожною действия / 3-карболинов пр норциональна числу оккупированных БДР. Так, / 3 - ССМ, оккупиру 40 % БДР нейронов гиппокампа, вызывает судороги, при 30 % эаняты рецепторов он обладает просудорожной активностью, а оккупация 5 БДР сопровождается облегчением обучения и памяти. Повторные инъекции FG 7142 ( по 40 мг / кг 3 раза за 6 дней) или норгармана ( 20 дней ежедневно) вызывают у мышей химическую раскачку проявляющуюся в том, что несудорожные дозы конвульсантов, в том числе FG 7142 или норгармана, способствуют возникновению клони-котонических судорог. Раскачка сопровождалась уменьшением свя зывания ( n ax) 35 S-бутилбициклофосфонотионата и снижением стимулированного мусцимоюм захвата 36С1 синаптосомами мозга, на основании чего сделано заключение, что просудорожное действи FC 7142 обусловлено ухудшением функционирования ГАМК-регули-руемых хлорных каналов [519,520] Однако, согласно [521], химический киндлинг посредством FG 7142 не сопровождается изменением количества и сродства БДР, а также способности ГАМК или диазепам усиливать связывания 3Н - лораэепама и 3S S грег-бутилбициклофосфо-нотионата. [46]
Вирус бешенства относится к РНК-содержащим вирусам рода Lyssavirus, семейства Rhabdoviridae. При бешенстве поражается центральная нервная система, поэтому диагностика заболевания основана на обнаружении в ткани мозга телец Бабеша-Негри ( в области гиппокампа), вирусного антигена или самого вируса. При исследовании животного, нанесшего укус, определяют наличие вируса или вирусного антигена в ткани слюнной железы с помощью РИФ и биологической пробы. [47]
Следует подчеркнуть, что это угасание реакций пирамидных нейронов гиппокампа селективно в отношении параметров стимула. Важным условием привыкания реакций пирамидных нейронов на приход сигналов от коры является потенциация синапсов нейронов зубчатой фасции, также приходящих на пирамидные нейроны гиппокампа. Таким образом, сигналы корковых детекторов поступают параллельно, прямо на пирамидные нейроны и через нейроны зубчатой фасции. При этом нейроны зубчатой фасции селективно блокируют локальные участки дендритов пирамидных нейронов, куда приходят корковые сигналы. Соседние синапсы корковых детекторов генерируют в дендритах пирамидных нейронов возбуждающие постсинаптические потенциалы ( ВПСП), которые развиваются без шунтирующего взаимодействия между участками мембраны, где они возникают. Гаммааминома-сляная кислота, ионофоретически локально аплицируемая на дендрит пирамидного нейрона, вызывает деполяризацию, которая связана с локальным повышением проводимости на данном участке мембраны дендрита. В результате соседние ВПСП подвергаются депрессии. ВПСП, вызванные на удаленных участках дендрита под влиянием вызванной гаммааминомасляной кислотой деполяризации, оказываются более эффективными в отношении генерации потенциалов действия. Этот механизм предположительно функционирует при депрессии корковых входов нейронами зубчатой фасции. [48]
Электроды обычно имплантировались в медиальное коленчатое тело, височную область коры, в ретикулярную формацию среднего мозга, образования переднего и заднего гипоталамуса, гиппокамп. [49]
Различные концентрации 3Н - лейцина ( 5 - 60 мкл, 1 мкКи / мкл) вводили в оба боковых желудочка мозга крыс. Взято было 20 животных весом 175 - 200 г. Крысу забивали через 1 5 часа после инъекции, извлекали мозг и вырезали из него два участка гиппокампа САЗ. Были выбраны именно эти участки, так как они связаны с изучавшейся поведенческой реакцией. [50]
Для изучения механизма образования условных рефлексов существенное значение имеет не только точная регистрация самой ответной реакции ( слюноотделения, движения и т.п.), но также исследование электрической активности, возникающей в различных мозговых структурах во время действия условного и безусловного раздражителей. Для регистрации электрической активности применяют электроды, хронически вживляемые в различные области или слои коры большого мозга, а также в специфические и неспецифические ядра таламуса, ретикулярную формацию, гиппокамп и другие отделы головного мозга. В опытах с условными рефлексами широко используются микроэлектродные методы, позволяющие регистрировать электрическую активность отдельных нейронов, участвующих в осуществлении условнорефлекторной реакции. Для автоматического анализа электроэнцефалограмм, регистрируемых с различных областей коры, в опытах на животных непосредственно во время условнорефлекторных реакций используют электронно-вычислительные машины. [51]
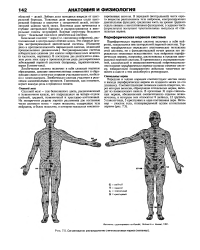 |
Сегментарное распределение спинномозговых нервов ( метамер. [52] |
Лимбическая система включает в себя сложные нервные образования, дающие многочисленные ответвления и образующие связи со многими старыми участками мозга, особенно с гипоталамусом. Лимбическая система участвует в регуляции эмоциональных процесов. Гиппокамп, как полагают, играет важную роль в процессах памяти. [53]
При действии раздражителя, обладающего определенным комплексом свойств, происходит селективное включение детекторов. При повторении раздражителя возбуждение детекторов остается постоянным. Однако влияние вклада этих детекторов в реакции нейронов гиппокампа ослабевает. Это ослабление селективно и ограничено именно теми детекторами, которые включались при действии раздражителя. При изменении стимула в реакцию вовлекаются ранее не действовавшие детекторы. Возбуждение этих детекторов достигает А - и Т - нейронов без ослабления. Это приводит к реакциям возбуждения в А-нейронах и реакциям торможения в Т - нейронах. Нейроны гиппокампа связаны с ретикулярной системой. Можно предположить, что А-нейроны связаны с активирующей системой, а Т - нейроны с инактивирующей системой мозга. [54]
 |
Модель коркового контроля над афферентными подавляющими и тормозными процессами ( в лобно-височную систему включается и миндалевидное ядро. [55] |
Этот более тонкий процесс относится к тем явлениям, которые математические психологи называют негативными случаями. Этот процесс следует за самой ранней фазой процесса регистрации и характеризуется началом активного игнорирования организмом тех явлений, которые уже происходили однажды, но теперь больше не подкрепляются. Имеются надежные доказательства того, что в этом процессе участвует гиппокамп, часть переднего мозга, примыкающая к миндалевидному ядру. Так, например, мы показали, что, если в ситуации различения число неподкрепляемых ( то есть негативных) стимулов варьирует от одного до четырех, выполнение задания нормальными животными почти не изменяется. [56]
Нарикашвили ( 1962), Н. Н. Зи-слина ( 1963), Л. А. Новикова ( 1963), Ю. С. Бородкина ( 1967), В. А. Крауз ( 1969) и др. убедительно показано, что мезэнцефалическая ретикулярная формация при пороговом раздражении повышает, а при надпороговой стимуляции подавляет активность коры больших полушарий. Она находится в реципрокных отношениях со структурами лимбической системы. Так, пороговое и надпороговое раздражение мезэнцефалической ретикулярной формации понижает активность гиппокампа; в свою очередь предварительная стимуляция этого образования блокирует активность восходящей ретикулярной формации. Тонус коры больших полушарий находится в значительной зависимости и от структур лимбической системы. [57]
В работе [98] также установлено, что наибольшая плотность участков, связывающих диазепам, находится в коре головного мозга крыс. Далее по убывающей плотности идут гипоталамус, мозжечок, средний мозг, гиппокамп, полосатое тело, продолговатый мозг и спинной мозг. [58]
Страницы: 1 2 3 4