Два - фермент
Cтраница 1
 |
Инициация репликации дромосомы Е. coli. [1] |
Два фермента обеспечивают высокую избирательность инициации синтеза ДНК, ограничивая инициацию репликации только ориджином. Это топоизо-мераза I и РНКаза Н, избирательно гидролизующая РНК в составе гибридных дуплексов с ДНК - Действие этих ферментов направлено против гибридных ДНК-РНК-участков, которые могут случайно образоваться на ДНК при транскрипции и послужить затравками для начала синтеза ДНК. [2]
 |
Биосинтез пиримидиновых нуклеотидов. [3] |
Два фермента, катализирующие последние две реакции, образуют другой комплекс и связаны также ковалентно. Вероятно, ко-валентное связывание функционально родственных ферментов и синтезирование в эквимолярных количествах является общим явлением у эукариот, что дает возможность создавать мультифер-ментные комплексы. [4]
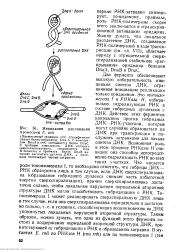 |
Инициация репликации хромосомы Е. coli. [5] |
Два фермента обеспечивают высокую избирательность инициации синтеза ДНК, ограничивая инициацию репликации только ориджином. Это топоизо-мераза I и РНКаза Н, избирательно гидролизующая РНК в составе гибридных дуплексов с ДНК - Действие этих ферментов направлено против гибридных ДНК-РНК-участков, которые могут случайно образоваться на ДНК при транскрипции и послужить затравками для начала синтеза ДНК. Возможная роль в этом процессе РНКазы Н очевидна: она способна непосредственно гидролизовать РНК во всех таких участках. Что касается роли топоизомеразы I, то необходимо отметить, что гибриды ДНК - РНК образуются лишь в том случае, если ДНК сверхспирализова-на ( образование гибридного дуплекса снимает часть избыточной энергии сверхспирализации), причем сверхспирализована достаточно сильно, чтобы локальные нарушения нормальной вторичной структуры ДНК могли способствовать гибридизации с РНК. [6]
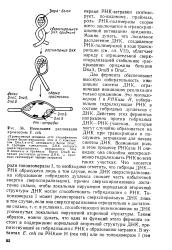 |
Инициация репликации. хромосомы Е. coli. [7] |
Два фермента обеспечивают высокую избирательность инициации синтеза ДНК, ограничивая инициацию репликации только ориджином. [8]
Эти два фермента отличаются не только характером действия на крахмал, но и различаются по своим свойствам. Ами-лаза кислотоустойчива, а а-амилаза в кислой среде быстро теряет свою активность; р-амилаза - термолабильна, а а-амилаза термостабильна и может действовать во время выпечки хлеба. [9]
У алкаптонуриков отсутствуют два фермента, участвующие в окислительном превращении тирозина: п-оксифенилпируватоксидаза и оксидаза гомогентизиновой кислоты. Первый фермент обнаружен только в печени, второй - - в почках. С стабилизируют эти ферменты и предохраняют их от инактивирования. [10]
Для рибулозобисфосфатного цикла характерны два фермента, не участвующие в других метаболических путях-фосфорибулокиназа и рибулозобисфосфат-карбоксилаза. Последний фермент представляет собой белок, в количественном отношении преобладающий над всеми другими белками на нашей планете. Рибулозобисфосфатный цикл-это восстановительный процесс, в котором СО2 восстанавливается до уровня углеводов. [11]
Основными ферментами неокислительной фазы являются два фермента: транскетолаза и трансальдолаза. Они отщепляют от фосфорилированных кетосахаров соответственно С2 - и С3 - фрагменты, перенося их на фосфоальдосахара. В результате взаимодействия фосфосахаров регулируется их количество в соответствии с потребностями клетки. [12]
Имеются данные, что в растениях встречаются два фермента, осуществляющие синтез полирибонуклеотидов. [13]
В большинстве животных и растительных клеток содержится два фермента, способных окислять () - изоцитрат - вещество, весьма распространенное в природе. [14]
Если идут два процесса, то должны существовать два фермента: ДНК - и РНК-полимеразы. Эти белки искали, и их действительно нашли в клетке. [15]
Страницы: 1 2 3 4