Дезаминирование
Cтраница 2
Дезаминирование проводят действием нитрита натрия на раствор полисахарида в уксусной кислоте при температурах от 0 до 50 С. [16]
Дезаминирование, например превращение А в Гк, вызывает еще не мутацию, а только премутацию. [17]
Дезаминирование аденина в составе полн-нуклеотида ( превращение в гипоксантин) меняет информац. Дезаминирование гуанина ( превращение его в ксантин) в составе матричных полинуклеотидов приводит к блокированию репликации и транскрипции. N-7 в составе матричных полинуклеотидов не сопровождается изменением генетич. [18]
 |
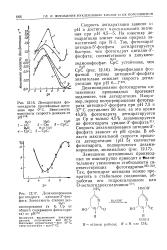
Константы скорости псевдопервого порядка при взаимодействии нуклеозидов с азотистой кислотой78. [19] |
Дезаминирование гуанозина под действием азотистой кислоты сопровождается побочными реакциями. [20]
 |
Дегидратация фотогидратов производных цито-зина при 0 С. Зависимость константы скорости реакции от рН 212.| Дезаминирование фотогидрата цитидин - З - фос-фата. Зависимость степени дез. [21] |
Дезаминирование фотогидратов ци-тозиновых производных протекает медленнее, чем дегидратация. Скорость дезаминирования цитидин-3 - фосфата зависит от рН среды. [22]
Дезаминирование дибензиламина дифторамином [ 9lJ приводит к дибензилу - типичному продукту превращения дибензил - аминонитрена. [23]
Дезаминирование цис - и / пранс-бицнкло - [3,1,0] - гексиламинов в водных растворах. [24]
Дезаминирование белков азотистой кислотой сопровождается вторичной реакцией, протекающей с участием фенольных групп и, возможно, также индольного и имидазольного колец. Тирозин диазотируется в о / 7го - положении, повидимому, путем образования нитрозопроизводных. Диазопроизводные белков, имеющие желтый или красноватый цвет, легко вступают в реакцию сочетания с фенольными производными, образуя окрашенные соединения. Эта реакция явно неспецифична, и, за исключением результатов, полученных при исследовании пепсина Филпотом и Смоллом [120], почти ничего не известно о природе и количестве реагирующих групп. Образующийся желтый диазопепсин обнаруживает максимум поглощения при 412 ту и обладает лишь 50 % активности исходного фермента. [25]
Дезаминирование гуанина ( XVI) в ксантин ( IV), протекающее при нагревании гуанина ( или п анин-сульфата) с нитритом натрия в кислой среде или при простом нагревании с концентрированной соляной кислотой, представляет интерес в качестве метода получения ксантина, поскольку исходный гуанин содержится в чешуе рыб, а также может быть выделен из нуклеиновых кислот. Превращению в ксантин может быть подвергнут непосредственно гидролизат нуклеиновых кислот или выделенный из него гуанозин. В последнем случае, наряду с ксантином ( IV), из маточников может быть выделена D-рибоза, являющаяся весьма ценным продуктом, используемым, например, в синтезе витамина В. [26]
Дезаминирование тирамина, норадренали-на, адреналина, алифатических моноаминов катализирует МАО; окисление гистамина и алифатических диаминов с короткой цепью углеродных атомов ( путресцин, кадаверин) катализирует ДАО. Ингибиторы МАО ( ипраниазид, гармин, паргилин) используют при лечении гипертонической болезни, депрессивных состояний. [27]
Дезаминирование алкилциклогэксиламинов обычно происходит с сохранением конфоркации. [28]
Дезаминирование дейтерированного п-нитроанилина проводили по методике, разработанной для легких веществ [7]; 13 8 г тонко измельченного н-нитроанилина смешивали с 90 мл ледяной уксусной кислоты и постепенно, за 30 мин. [29]
Дезаминирование адениловой кислоты является, таким образом, процессом, направленным на устранение избытка этого вещества в мышечной ткани, и, по-видимому, прямо не связано с актом сокращения мышечного волокна. [30]
Страницы: 1 2 3 4