Окислительное дезаминирование
Cтраница 1
Окислительное дезаминирование превалирует у животных, растений и многих видов микроорганизмов, другие типы дез а минирования встречаются главшлм образом у некоторых анаэробных микробов. Окислительное дел ям и ни ров анис протекает тн два этапа. [1]
Окислительное дезаминирование доказано для животных, растений и большинства микробов. [2]
Окислительное дезаминирование а-амино-кислот в а-кетокислоты и моноаминов в альдегиды. Окислительное дезаминирование а-аминокислот катализируют флавопротеиды-оксидазы L-аминокислот, катализирующие окисление природных аминокислот L-ряда, и оксидазы D-аминокислот, действующие на неприродные аминокислоты D-ряда. [3]
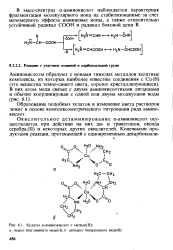 |
Хелаты а-аминокислот с медью ( Н. [4] |
Окислительное дезаминирование а-аминокислот осуществляется при действии на них ди - и трикетонов, оксида серебра ( II) и некоторых других окислителей. [5]
Окислительное дезаминирование ос-аминокислот под действием 1 2 3-ин-дантриона - нингидрина - лежит в основе их качественного обнаружения. [6]
Реакции окислительного дезаминирования катализируются ферментами, которые получили название оксидаз аминокислот. Они представляют собой двухкомпонентные ферменты, в состав активной группы которых входит флавинаденин-динуклеотид. Эти ферменты характеризуются высокой специфичностью. [7]
Механизм окислительного дезаминирования моноаминов является необратимым и протекает в две стадии. [8]
На примере окислительного дезаминирования впервые установлено и стереохимическое течение перегруппировки пинаколинового типа. [9]
В результате окислительного дезаминирования образуется аммиак, который идет на синтез мочевины. Основными конечными продуктами обмена белков являются мочевина, вода и двуокись углерода. [10]
Первая стадия окислительного дезаминирования - ферментативная, а вторая - гидролиз иминокислоты - протекает самопроизвольно. [11]
Данные ферменты катализируют окислительное дезаминирование катехоламинов. Биохимиками более лзучена структура и функция моноаминоксидазы, которая в отличие от холинэстеразы является сложным металло-протеидом, имеющим в составе своей молекулы активную каталитическую часть особой химической структуры, именуемую в биохимии простетической, или коферментной в включающую пиридоксальфосфатную группировку и атомы меди. В свою очередь пиридоксальфосфат состоит из пиридоксина ( витамина В6) и фосфорной кислоты. Согласно принятым данным 22 именно пиридоксин и медь являются важнейшими компонентами активных каталитических центров моноаминоксидазы и ряда других ферментов; получивших общее название пиридоксале-вых. [12]
Специфическая D-аспартатоксидаза катализирует окислительное дезаминирование D-аспарагиновой кислоты в щавелевоуксусную кислоту. Природа флавиновой простетической группы фермента не установлена. [13]
Аммиак образуется при окислительном дезаминировании и аэробном декарбоксилировании аминокислот. Аммиак очень токсичен, так как он нарушает кислотно-основное равновесие. В печени происходит ряд реакций, в результате которых аммиак удаляется из организма. В цикле орнитин реагирует с двуокисью углерода и аммиаком с образованием цитруллина. Цитруллин превращается в аргининянтарную кислоту и затем в аргинин, который при гидролизе дает исходный орнитин и мочевину. Мочевина поступает с током крови в почки и уносится из организма с мочой, а орнитин возвращается в цикл. При декарбоксилировании глутаминовая кислота в печени реагирует с аммиаком и образует глутамин, который током крови переносится в почки, где он гидролизуется, давая аммиак и глутаминовую кислоту. [14]
При действии нингидрина происходит окислительное дезаминирование а-аминокислот. Выделяющийся при этом аммиак реагирует с нингидрином, образуя продукт, окрашенный в голубой цвет. Пролин и оксипролин в тех же условиях дают красную окраску. [15]
Страницы: 1 2 3 4