Длина - хромосома
Cтраница 2
ГЕНЫ, по хромосомной теории наследственности, дискретные единицы, на к-рые дифференцированы по своей длине хромосомы ( элементы ядра животных и растит, клеток) и посредством к-рых передаются по наследству признаки и свойства организма. [16]
В результате поиска условно-летальных температурочувствительных мутантов бактериофага Т4 были обнаружены сотни различных мутантов с беспорядочным распределением мутаций по всей длине фаговой хромосомы. Так, оказалось, что продукт гена 42 представляет собой специфический фермент, необходимый для синтеза оксиметил - dCMP ( гл. Гены 20 - 24 в числе прочих, по-видимому, кодируют синтез белков головки фага, так как при мутациях этих генов образуются ( Нормальные отростки, но белки головки не синтезируются. Ген 23 кодирует, по-видимому, синтез основных субъединиц головки, а ген 20 как-то связан с процессом образования колпачка ( capping) на конце головки. Эти мутанты образуют цилиндрические полиголовки вместо обычных головок. У мутантов с измененными генами 25 - 29 нарушено строение базальной пластинки и совсем не образуется отросток, а у мутантов с измененными генами 34 - 38 отсутствуют нити отростка. [17]
Принято различать крупные и мелкие хромосомные перестройки; к последним относятся такие инверсии, дупликации, нехватки или транслокации, которыми захвачены только маленькие участки по длине хромосомы. Маленькие дупликации и гетерозиготные нехватки у дрозофилы обычно жизнеспособны, в противоположность крупным, нежизнеспособным. Так, нехватка части района 1А ( по карте хромосом слюнных желез Drosophila melanogaster) ведет себя как рецессивная видимая мутация:; в гомозиготном состоянии она вызывает появление у мухи желтой окраски тела вместо нормальной серой. Нехватка, затрагивающая диск ЗС7, действует как доминантная мутация, вызывая в гетерозиготном состоянии выемки на крыльях. В гомозиготном состоянии она летальна. [18]
Исходя из первой интерпретации, мы используем экспериментальные данные, чтобы определить число генов, способных дать летальные мутации; исходя из второй интерпретации, мы определяем длину хромосомы, в пределах которой они локализованы; эти две величины, несомненно, связаны между собой. [19]
V - размер ( число хромосом) популяции; Q - множество номеров членов популяции, в хромосомах которых имеется схема Н; ДН) - длина схемы Н, т.е. расстояние ( число позиций) между крайними локусами схемы, так, в случае (2.5) имеем ДН) 8 - 4 4; ДН) - порядок схемы Н ( число локусов в Н), в примере (2.5) порядок равен 3; п - длина хромосомы. [20]
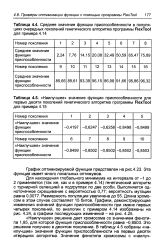
Для нахождения глобального минимума на интервале от - 1 до 2 применяется ( так же, как и в примере 4.14) генетический алгоритм с турнирной селекцией в подгруппах по две особи. Длина хромосом в этом случае составляет 15 битов. [22]
Применяется генетический алгоритм с турнирной селекцией в подгруппах по две особи с одной точкой скрещивания. Длина хромосом для решаемой задачи равна 99 битам - по 11 генов на каждую переменную. [23]
Второй прием, позволяющий уменьшить длину хромосомы, основывается на том, что если цепи не имеют горизонтальных ограничений, то их взаимное положение либо не важно, либо определяется из соотношений с остальными цепями. Следовательно, длина хромосомы будет равна L NGO - NRVO, где NGO - число горизонтальных ограничений, NRVO - число вертикальных ограничений в расширенном графе вертикальных ограничений. [24]
Как видно, длина хромосомы получается достаточно большой ( для примера на рис. 6.33 она равна 15), что не приемлемо. Поэтому для уменьшения длины хромосомы используется анализ расширенного графа вертикальных ограничений и графа горизонтальных ограничений. [25]
Аналогично, расширение области определения переменной х также потребует применения более длинных хромосом. Из этих наблюдений можно сделать вывод, что длина хромосом зависит от ширины области определения х и от требуемой точности решения. [26]
Применяется генетический алгоритм с турнирной селекцией в подгруппах по две особи с одной или двумя точками скрещивания, а также с селекцией по методу рулетки с одной точкой скрещивания. В этом случае с учетом меньшей требуемой точности длина хромосом составляет 22 бита - по 11 генов на каждую переменную. [27]
ДНК увеличиваются, что и приводит к увеличению диаметра н длины хромосом. Благодаря неравномерной спирализации в политенных хромосомах образуются диски ( темные поперечные полосы), выявляемые при окраске хромосом. Число, размер и характер расположения дисков специфичны для вида. [28]
Об одном из гипотетических механизмов разлома многонитчатой хромосомы говорил Спитковский. Это - разрушение слабых взаимодействий между концами молекул ДНК: известно, что по длине хромосомы должно быть значительное число концевых стыков молекул ДНК с молекулярным весом порядка 107, а может быть и некоторое перекрывание концов молекул ДНК. В таком случае должны существовать преимущественные места разломов по длине хромосомы и мы знаем, что такие места действительно есть. [29]
В настоящее время он популярен для решения различных задач теории расписаний. Вместо использования разрезающей точки ( точек) здесь определяют двоичную маску, длина которой равна длине заданных хромосом. [30]
Страницы: 1 2 3 4