Длина - хромосома
Cтраница 3
Популяция представляет собой множество pi, состоящее из хромосом, где Np - численность популяции. Геном pi - цепочка из L символов, pi L, то есть, L - длина хромосомы. Вводится ограничение: L и Np не изменяются в процессе эволюции. [31]
При последующем же удвоении, протекавшем в отсутствие Н3 - тимидина, метка включалась только в одну из каждой пары дочерних хромосом. Из этих результатов следует, что ДНК синтезируется в виде целой нити, которая тянется по всей длине хромосомы, и что при последующих репликациях ее целостность не нарушается ( ср. Мезельсона и Сталя, стр. Кроме того, эти данные говорят и о том, что в ходе митоза вновь синтезированная ДНК распределяется между дочерними хромосомами поровну. [32]
В настоящее время для обнаружения новых мутаций, возникающих в Х - хромосоме дрозофилы, применяют другой метод, более простой и эффективный, чем метод С / В. В так называемой линии Меллер-5 Х - хромосома содержит две инверсии, которые препятствуют перекресту по всей длине хромосомы. [33]
Таким образом, в реплицирующих участках хромосомы от 1 р может возникнуть 14 пар ионов. Но такие небольшие дозы облучения, как известно, не могут вызвать серьезных повреждений, так как эти 14 ионизации будут распределены не локально, а дискретно по всей длине хромосомы, по числу реплицирующих участков ( репликонов), в которых одновременно может идти синтез ДНК. Биологический эффект ( гибель клеток или разрыв хромосом) наблюдается при более высоких дозах облучения - порядка нескольких десятков рентгенов. [34]
Выбор исходной популяции связан с представлением параметров задачи в форме хромосом, т.е. с так называемым хромосомным представлением. Это представление определяется способом кодирования. Длина хромосом зависит от условий задачи, точнее говоря - от количества точек в пространстве поиска. [35]
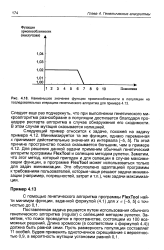 |
Наименьшее значение функции приспособленности в популяции на последовательных итерациях генетического алгоритма для примера. [36] |
Поставленная задача решается путем использования обычного генетического алгоритма ( regular) с селекцией методом рулетки. Заметим, что поиск проводится в пространстве, состоящем из 100 возможных решений. Длина хромосом ( в соответствии с формулой 4.4)) должна быть равной семи. Пусть размерность популяции составляет 11 особей. [37]
В нашем описании мейоза было указано, что сильное укорочение хромосом в период между началом профазы и первой метафазой вызвано спиральным закручиванием хромосомной нити, в результате которого она постепенно уплотняется. Известно также, что обычные хромосомы имеют строение спирали, которое можно выявить, воздействуя на них определенными химическими веществами. Поэтому имеются основания полагать, что огромная длина хромосом слюнных желез обусловлена, по крайней мере частично, развертыванием хромосомной спирали. [38]
Об одном из гипотетических механизмов разлома многонитчатой хромосомы говорил Спитковский. Это - разрушение слабых взаимодействий между концами молекул ДНК: известно, что по длине хромосомы должно быть значительное число концевых стыков молекул ДНК с молекулярным весом порядка 107, а может быть и некоторое перекрывание концов молекул ДНК. В таком случае должны существовать преимущественные места разломов по длине хромосомы и мы знаем, что такие места действительно есть. [39]
Показано, что у трипаносом концы хромосом удлиняются при каждом раунде репликации, но время от времени происходит их существенное укорочение. При размножении дрожжей хромосомы не растут, но в одном из дрожжевых мутантов с нарушением нормального клеточного цикла на непермиссивной температуре начинается удлинение теломер. Можно думать, что ген, маркированный этой мутацией, участвует в ограничении длины хромосом. [40]
Позвольте сказать несколько слов по поводу доклада Струч-кова. В этом интересном сообщении сделана, по сути дела, попытка ответить на один из наиболее важных вопросов радиобиологии: в каком состоянии находится ДНК в клетке и в связи с этим - в чем заключается повреждение этих генетически важных макромолекул. Я не могу согласиться с точкой зрения, что хромосома состоит из большого числа нитей ДНК, непрерывных по всей длине хромосомы. Из теории кинетики деградации хорошо известно, что в этом случае доза ионизирующей радиации, которая привела бы к разрыву хромосомы, имела бы величину, на много порядков большую, чем это имеет место на самом деле. [41]
 |
Частоты аллелей и генотипов в большой популяции со случайным скрещиванием1. [42] |
Для возникновения аллелей достаточно, чтобы два гомологичных гена различались всего одним нуклеотидом. Во многих случаях замена одного нуклеотида приводит к значительным различиям между продуктом измененного гена и нормальным белком. Однако множество однонук-леотидных замен не приводит к синтезу измененных генных продуктов, а кроме того, замены могут происходить в некодирующих областях ДНК и не приводить ни к каким последствиям. Такие безвредные замены, распределяясь по всей длине хромосомы, порождают полиморфные сайты ( маркерные локусы, генетические маркеры), которые можно использовать для генетического картирования. Но сначала эти полиморфные сайты нужно обнаружить. [43]
На международном генетическом конгрессе, происходившем в 1927 г. в Берлине, Меллер смог сообщить, что он определил частоту спонтанных мутаций в Х - хромосоме плодовой мушки и что после облучения частота этих мутаций очень сильно увеличилась. Это увеличение частоты мутаций в несколько сот, а то и тысяч раз зависело от дозы облучения, а в некоторых опытах практически у каждой особи было обнаружено по одному или по нескольку вновь возникших му-тантных признаков. Меллер сообщил также, что экспериментально индуцированные мутации были аналогичны спонтанным и что некоторые из индуцированных мутаций, как показал генетический анализ, оказались либо идентичными спонтанным мутациям, либо представляли собой новые аллели уже известных ранее локусов. Далее было установлено, что индуцированные мутации распределены по всей длине хромосом и возникают в Х - хромосоме с такой же частотой ( в расчете на единицу длины), как и в аутосомах. [44]
Карта Х - хромосомы из слюнных желез разделена на 120 приблизительно одинаковых отрезков, обозначенных буквами. Последние шесть отрезков представляют проксимальный гетерохроматин. Имеются данные, свидетельствующие о том, что эти участки содержат гетерохроматин. Мы приходим к заключению, что у дрозофилы разрывы расположены довольно равномерно по длине хромосомы, и если учитывать длину митотических хромосом, а не хромосом слюнных желез, то разрывы в эухроматиновых и гетерохроматиновых районах происходят примерно с одинаковой частотой. [45]
Страницы: 1 2 3 4