Оротовая кислота
Cтраница 2
В работе [484] излагается потенциометрический метод титрования оротовой кислоты и смесей ее с тио -, нитро - и другими производными в среде ДМСО, в котором анализируемые соединена достаточно хорошо растворяются. Найдено, что лучшим титранток является изопропаноловый раствор гидроксида тетраэтиламмония Кривые титрования смесей в зависимости от титруемых объекте. [16]
Известно, что это соединение является предшественником оротовой кислоты ( стр. [17]
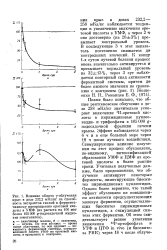
Кл / кг наблюдается тенденция к увеличению включения оротовой кислоты в УМФ, а через 2 т оно достоверно ( на 20 3 %) превышает контрольный уровень. В последующие 3 ч этот показатель постепенно снижается до контрольных значений. Позже было показано, что общее рентгеновское облучение в дозе 258 мКл / кг значительно ускоряет включение 3Н - оротовой кислоты в пиримидиновые нуклео-зидди - и трифосфаты в 105 000 g - надосадочной фракции печени: крысы. Эффект наблюдается через 6 ч и в еще большей мере через 18 ч после лучевого воздействия. Стимулирующее влияние излучения на этот процесс обусловлено, по-видимому, интенсификацией: образования УМФ и ЦМФ из оротовой кислоты в более ранние-сроки. Учитывая полученные данные, было предположено, что облучение активирует некоторые-ферменты, лимитирующие биосинтез пиримидиновых нуклеотидов. Однако более вероятно, что такой эффект обусловлен не повышением активности предсуществующих молекул ферментов, осуществляющих биосинтез пиримидиновых нуклеотидов, а стимуляцией синтеза этих ферментов. [19]
Основной путь синтеза пиримидинов de novo включает образование оротовой кислоты из простых предшественников. Аспара-гиновая кислота карбамилируется карбамилфосфатом ( образующимся из аммиака, двуокиси углерода и АТФ) с образованием карб-амил - Ь - аспарагиновой кислоты ( уреидоянтарной кислоты), которая при циклизации превращается в дигидрооротовую кислоту. Ферментативное окисление дает оротовую кислоту, которая с рибо-зо-1 - пирофосфат-5 - фосфатом образует оротидин-5 - фосфат. Последний при действии декарбоксилазы превращается в уридин-5 - фосфат ( см. схему на стр. [20]
Влияние оротовой кислоты и витамина А на включение 214С - оротовой кислоты в кислотораство-ри. [21]
Однако в общем случае пути, не включающие промежуточного образования оротовой кислоты, имеют большее значение в катаболических процессах. [22]
В частности, было обнаружено 2 - 4-кратное повышение скорости включения меченой оротовой кислоты в РНК ободочной кишки крыс с гиповитаминозом А после предварительного введения им витамина А. Однако подобный эффект не был выявлен в тонком кишечнике. [23]
Через 2 часа раствор подкисляют концентрированной соляной кислотой и выпавшую в осадок оротовую кислоту собирают и перекристаллизовывают из воды. [24]
Показано, что для урацила и других оснований протекают иные ферментативные синтезы, аналогичные гликозилированию оротовой кислоты рибозо-1 - пирофосфат-5 - фосфатом. Последовательное действие пиримидиннуклеозидфосфорилазы ( которая также осуществляет трансгликозилирование вследствие обратимого характера реакции) и пиримидиннуклеозидкиназы обеспечивает процесс, который протекает в определенных условиях при синтезе нуклео-тидов. [25]
Указанное предположение подтверждается и тем, что добавление актиномицина D к 105 000 g - надосадочной фракции печени не влияет на ферментативное превращение оротовой кислоты в УМФ [ Ghetty К. [26]
Показано, что через 3 ч после общего уоблучения крыс в дозе 1132 мКл / кг образование УМФ de novo в печени из 6 - 14С - оротовой кислоты при 40-минутном ее включении in vivo остается на контрольном уровне. Можно предположить, что в ранние сроки после облучения животных в сверхлетальных дозах эффект радиационной стимуляции превращения оротовой кислоты в УМФ в этой ткани уже не проявляется. Однако через 12 ч после облучения некоторая тенденция к активации еще сохраняется. По-видимому, при действии сверхлетальных доз элиминируется активация в печени синтеза оротат-фосфорибозилтрансферазы и ( или) оротидин-5 - фосфат-де-карбоксилазы, катализирующих двухэтапный процесс образования УМФ из оротовой кислоты. GI, когда происходит биосинтез ферментов синтеза предшественников нуклеиновых кислот. [27]
Возможно существование и другого пути биосинтеза пирими-динов, проходящего через карбомоил - 3-аланин, соответствующие рибонуклеозид и рибонуклеотид и затем дигидроуридин-5 - фосфат [59]; в этом случае оротовая кислота не принимает участия в образовании пиримидинового кольца. [28]
По полученным нами данным, через 15 мин после у-рблучения крыс в дозах 232 2 - 258 мКл / кг способность экстрактов вилочковой железы к биосинтезу УМФ из оротовой кислоты еше не нарушается. К концу 1-го часа после облучения наблюдается тенденция к усилению конечного двухэтапного биосинтеза de novo УМФ. В последующие сроки включение оротовой кислоты в УМФ постепенно снижается. Уже через 2 ч после облучения животных этот процесс угнетается на 20 %, через 5 ч - на 40 12 %, а через 6 / 2 ч - на 50 % контрольного уровня. В селезенке крыс в течение первых 60 мин после облучения этот процесс практически не нарушается. На протяжении следующих 3 ч процесс ферментативного превращения оротовой кислоты в УМФ сохраняется на этом уровне. [29]

Страницы: 1 2 3 4