Компонент - мембрана
Cтраница 2
В этом случае может быть понятна большая глубина наблюдаемых изменений подвижности зонда, связанного с белковой компонентой мембран, по сравнению с глубиной изменения подвижности зонда, локализованного в ли-пидной области мембраны, а также понятен температурный сдвиг начала структурных перестроек, последовательно захватывающих оба компонента мембраны. [16]
В связи с тем что влияние солевого фона на коэффициент активности ионов, определяемых в природных пресных водах, элиминируется фоновым электролитом, обеспечивающим постоянный коэффициент активности ( ц 1), присутствие посторонних солей может сказаться на показании электродов лишь в связи с химическим взаимодействием их с галогенидами или с компонентами мембраны электрода. Поэтому макрокомпоненты природных вод ( Na, K Mg ( II), HCOjj, SO j -, H2SO3, NOg -) не влияют на показания галогенидных электродов. [17]
Обобщая имеющиеся в литературе сведения о полиенрези-стентных ( пол - Р) штаммах грибов, а также об одноклеточной зеленой водоросли - хламидомонады, А. Б. Левченко и др. ( 1976) отмечают, что у этих организмов есть значительное сходство в генетико-биохимических механизмах пол - Р, связанных с качественными и ( или) количественными изменениями стерольных компонентов мембран. Сходство наблюдается при теистической детерминации признака пол - Р, в характере зависимости экспрессивности мутантных пол - Р генов от действия факторов внешней среды и генетического фона, в особенностях плейотропных эффектов полиенрезистентных генов. [18]
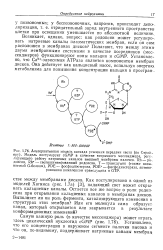
Возникает, однако, вопрос: как родопсин может регулировать натриевые каналы плазматических мембран, если он сам: расположен в мембранах дисков. АТРаза является компонентом мембран дисков. [20]
Более точным было бы предположение о существовании сильного притяжения подобной мембраны к поверхности кремнезема, которое, вероятно, обусловливается присутствием фос-фатидилхолина. Этот тип соединения представляет собой компонент мембран в большинстве высших организмов и характеризуется наличием группы, содержащей четвертичный аммониевый ион. [21]
Конечно, остается возможность, что этот липид каким-то образом переносится в митохондрии и там соединяется со структурным белком. Таким образом, проблема переноса и сборки компонентов мембран с образованием функционирующих клеточных структур остается загадкой, разрешить которую предстоит в будущем. [22]
Помимо исследования специфического взаимодействия белковых и липидных компонент мембраны, проявляющегося в процессах рецепции, метод спинового зонда используется и для изучения достаточно общих закономерностей липид-белковых взаимодействий. Именно благодаря влиянию белков на состояние липидных областей мембран жирорастворимые зонды позволяют следить за состоянием белковых компонент мембраны. Так, в работе [188] при исследовании температурной зависимости подвижности зонда СП ( 5, 10) в мембранах саркоплазмати-ческого ретикулума и в работах [189] при исследовании температурной зависимости подвижности зонда АХЩ14) в мембранах бактерий Micrococus lysodeikticus, наряду с обычными структурными переходами в липидных областях мембраны, обусловленных самими липидами, обнаружены структурные переходы в липидных областях мембраны, которые исчезали при тепловой денатурации мембранных белков, что свидетельствует об индукции этих переходов конформационными превращениями мембранных белков. [23]
Хигучи и др. [351] установили, что пластифицированные мембранные электроды обладают высокой специфичностью по отношению к относительно гидрофобным органическим катионам и анионам. Авторы [351] полагают, что некоторые органические пластифицированные матрицы с ограниченно гидрофильным характером можно использовать в качестве гелеобразующего компонента мембраны. Выбор конкретной матрицы зависит прежде всего от ее совместимости с желаемыми жидкими пластифицирующими компонентами. Эти жидкие компоненты подбираются по их способности сольватировать интересующие аналитика ионы. Для того чтобы электрод обладал высокой специфичностью, необходимы жидкие растворители с особенно высокой степенью специфичности в отношении образования соответствующих сольватов. Время отклика электрода достаточно мало; в растворах с концентрацией более 10 5 М равновесие достигается менее чем за 1 мин. Электрод с поливинилхлоридной ( ПВХ) мембраной, пластифицированной М М - диметилолеоамидом ( халькомид 18 - OL), имеет нернстову электродную функцию относительно ионов тетрабутиламмония. Электрод с такой же мембраной, как показано на рис, 9.1, характеризуется линейной электродной функцией для анионов тетрафенилбората только в области высоких концентраций. [24]
Полистирол и полиметилметакрилат, взятые в качестве связующего материала, не обладают достаточной эластичностью, вследствие чего при набухании ионитового компонента мембраны неизбежно происходит нарушение сплошной структуры пленки. [25]
Каковы особенности белков, связанных с мембранами, у организмов, адаптированных к давлению: не отличаются ли их структура и функции особой чувствительностью к этому фактору. Различны ли фосфолипидные компоненты мембран у организмов, живущих на разных глубинах. [26]
Сингер и Никольсон ( 1972) предложили жидкостно-мозаичную модель строения мембраны: мембрана представляет собой липидный бислой, в котором расположены глобулярные белки. По этой модели мембрана представляет собой динамическую систему, в которой молекулы белка относительно свободно плавают в липидном море в виде айсбергов. Кроме того, компоненты мембраны с внутренней стороны связаны с системой белков цитокаркаса. Согласно решетчато-мозаичной модели мембрана представляет собой липидный бислой, включающий молекулы белков, заключенные в ажурный каркас. Молекулы белков, которые связаны с цитокаркасом, малоподвижны; остальные могут относительно свободно перемещаться в плоскости мембраны. Так, в мембране эритроцитов интегральный анионпереносящий белок связан с белками цитокаркаса - спектрином, анкирином. При гемолизе эти связи нарушаются. [27]
 |
Натриевые каналы в возбудимых мембранах. [28] |
При выделении компонентов ионного канала подобный контроль не подходит. При этом следует учесть тот факт, что выделенный материал должен содержать все функционально важные компоненты канала. В связи с тем что эксперименты по реконструкции компонентов нервных мембран оказались не достаточно перспективными, а также весьма сложными, возникла необходимость разработки новых методов. Предварительно электрофизиологическими методами было показано их селективное взаимодействие с определенным каналом. В случае натриевого канала использовались меченные тритием тетродотоксин [ 3Н ] ТТХ или сакситок-син [ 3H ] STX [13], так как их константы диссоциации ( 10 - 9М) позволяют проводить радиолигандный анализ. [29]
Периферические нервные волокна высших животных часто окружены небольшим слоем клеток сателитов, которые называются шванновскими клетками. Шванновские клетки частично обволакивают нервное волокно и выполняют функцию защитного покрытия. По-видимому, шванновски клетки участвуют в синтезе ферментов и компонентов мембран нервных волокон. Большинство нервных волокон позвоночных животных, кроме самых тонких, окружены также толстой неклеточной жировой оболочкой, которую называют миелиновой оболочкой. Миелиновая оболочка порождается шванновскими клетками. В виде многих слоев она оборачивается вокруг нервного волокна. [30]
Страницы: 1 2 3