Компонент - дыхательная цепь
Cтраница 2
При этом через два образца пропускают два луча одной и той же длины волны. Допустим, что у одного образца компоненты дыхательной цепи окислены вследствие отсутствия субстрата. У другого образца в анаэробных условиях в присутствии субстрата компоненты дыхательной цепи находятся в восстановленном состоянии. Разница в оптической плотности двух образцов, определенная при нескольких длинах волн, позволяет получить разностный спектр. [16]

Отданные субстратами восстановительные эквиваленты ( протоны и электроны) переносятся на плазматическую мембрану или на внутреннюю мембрану митохондрий. Этот перепад заряда возникает благодаря определенному расположению компонентов дыхательной цепи в мембране. [18]
Кроме перечисленных выше компонентов, с переносом водорода связаны также кофермент Q, а-токоферол и витамин К. Однако имеющиеся данные недостаточны, чтобы считать эти соединения компонентами дыхательной цепи. [19]
Убихиноны переносят электроны через липидные слои мембран. Они играют важную роль в процессах тканевого дыхания, являясь компонентами дыхательной цепи ( гл. [20]
Цитохромы в высших растениях содержатся в значительно меньшей концентрации, чем в животных, дрожжах и бактериях. Тем не менее все приведенные в табл. 19 цитохромы, являющиеся компонентами дыхательной цепи, возможно за исключением цито-хрома Ci, найдены в растениях. [21]
Роль кофермента Q в дыхательной цепи у животных пока еще не выяснена. Грин и его сотрудники [15] считают, что кофермент Q представляет собой компонент дыхательной цепи и функционирует между цитохромом b и участком, чувствительным к антими-цину ( см. стр. [22]
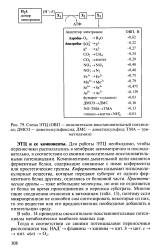 |
Схема ЭТЦ ( ОВП - окислительно-восстановительный потенциал. ДМСО - диметилсульфоксид. ДМС - диметилсульфид. ТМА - три. [23] |
Для работы ЭТЦ необходимо, чтобы переносчики располагались в мембране асимметрично и последовательно, в соответствии со своими окислительно-восстановительными потенциалами. Компонентами дыхательной цепи являются ферментные белки, содержащие связанные с ними коферменты или простетические группы. Коферментами называют низкомолекулярные вещества, которые передают субстрат от одного ферментного белка другому, отделяясь от белковой части. Простетические группы - тоже небольшие молекулы, но они не отделяются от белка во время присоединения и переноса субстрата. Многие такие соединения относятся к витаминам ( табл. 13), поэтому если микроорганизм не способен сам синтезировать некоторые из них, то это вещество или его предшественник необходимо добавлять в питательную среду. [24]
Большинство субстратов поступает в цепь при участии связанных с пиридиннуклео-тидами дегидрогеназ. В вопросе о точной природе компонентов дыхательной цепи п их последовательности существуют разногласия. Последовательность компонентов, показанная на фиг. [25]
Регенерация АТР при фосфо-рилировании в дыхательной цепи и фотосинтетическом фосфорилирова-нии протекает в мембранах. АТР-синтаза, так же как и компоненты дыхательной цепи, является составной частью мембраны. Каким образом происходящий в дыхательной цепи перенос водорода и эдйстронов сопряжен с синтезом АТР, до конца еще не выяснено. Однако многочисленные эксперименты показали, что регенерация АТР происходит только в пространствах, окруженных со всех сторон мембранами-в пузырьках, или везикулах. Процессы переноса водорода и электронов теснейшим образом сопряжены с перемещением протонов, а этот процесс в свою очередь необходим для регенерации АТР. [26]
В основе циклической структуры обоих витаминов лежит кольцо 1 4-нафтохинона. Заметим, что животные ткани наделены способностью синтеза боковых изопреновых цепей, но не могут синтезировать нафтохиноновый компонент. У большинства бактерий витамин К является компонентом дыхательной цепи вместо убихинона. [27]
Интактные митохондрии представляют собой осмотически активные пузырьки, отделенные от гиалоплазмы ( или среды инкубации при эксперименте in vitro) двумя мембранами. Таким образом, существуют четыре топологически различных пространства: внешняя мембрана, межмембранное пространство, внутренняя мембрана и внутреннее пространство - матрикс. Ферменты цикла трикарбоновых кислот сосредоточены в матриксе; компоненты дыхательной цепи, транслоказа адениннуклеотидов и АТФ-синтетазный комплекс прочно связаны с внутренней мембраной, в межмембранном пространстве локализована аденилаткиназа, а во внешней мембране - моноаминооксидаза. [28]
При этом через два образца пропускают два луча одной и той же длины волны. Допустим, что у одного образца компоненты дыхательной цепи окислены вследствие отсутствия субстрата. У другого образца в анаэробных условиях в присутствии субстрата компоненты дыхательной цепи находятся в восстановленном состоянии. Разница в оптической плотности двух образцов, определенная при нескольких длинах волн, позволяет получить разностный спектр. [29]

Страницы: 1 2 3