Липкий конец
Cтраница 1
Липкие концы ( Cohesive ends) Взаимно комплементарные одноцепочные участки ДНК, выступающие по концам двухцепочечной молекулы; образуются в результате ступенчатых разрезов двухцепочечных ДНК. [1]
Липкие концы - особенность структурной организации ДНК, которая хорошо нл-люстрир уется таким примером. При нагревании ДНК бактериофага Я, до 65 С и последующем медленном охлаждении образуются молекулы с коэффициентом седиментации 37 S. Быстрое же охлаждение ведет к образованию молекул с коэффициентом седиментации 32 S. Это объясняется тем, что 5 -концы ДНК выступают за пределы комплементарных З - концов в виде взаимно комплементарных одиночных цепей. Основания липких концов спариваются, и линейная молекула ДНК превращается в кольцевую, которая при 65 С разрывается. Установлено, что липкие концы представлены последовательностью, состоящей из 12 мономерных звеньев. Результаты электрон-вой микроскопии показывают, что 37 S-форма ДНК является кольцевой, а 32 S-форма - линейной. Таким строением характеризуются ДНК лизогениаирующих бактериофагов. [2]
Если у гена липкие концы нужного строения отсутствуют, то. Удобным для этой цели ферментом является терминальная дезоксинуклеотидил трансфераза ( выделяемая из вилочковой железы телят), которая катализирует безматричный перепое дезоксинуклеотидных остатков от дезоксинуклеозид-5 - трифосфатов на 3 -концевую гндроксигруппу олиго - или полидезоксинуклеотидов. Слипание гена и плазмиды происходит за счет комплементарных олигодезоксигуани ловых и олигодезокси-цитидиловых последовательностей, причем образуется кольцевая структура с брешами в каждой нити. После этого пики ликвидируются с помощью ДНК-лигазы. [3]
Многие рестрикционные эндонуклеазы, расщепляя ДНК, дают липкие концы ( см. с. Различные ДНК могут быть соединены при помощи ДНК-лигазы и по тупым концам. [4]
Прн работе с рекомбинантнымн молекулами ДНК возникает необходимость замены одного типа липких концов на другой. Для этой цели используются адапторные линкеры - частично двухце-почечные нуклеотиды, содержащие липкие концы, соответствующие двум разным рестриктазам. Третий тип линкеров позволяет перейти от тупого конца ДНК к липкому, и их можно назвать дополняющими. [5]
Один из подходов состоит в использовании рестриктирующих эндонуклеаз для получения фрагментов ДНК с липкими концами. [6]
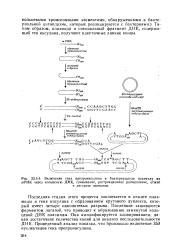 |
Включение гена препроинсулина в бактериальную плазмиду из мРНК через кольцевую ДНК, связывание, рестрикционное расщепление, отжиг. [7] |
Таким образом, плазмида и специальный фрагмент ДНК, содержащий ген инсулина, получают идентичные липкие концы. [8]
 |
Схема терминации синтеза ( цепи фага рХ174. [9] |
После попадания в клетку линейной двухнитевой ДНК фага X она циркуляризуется за счет взаимодействия между липкими концами и превращается в ковалентно-непрерывную сверхспирализо-ванную форму. [10]
 |
Схема терминации синтеза ( цепи фага рХ174. [11] |
После попадания в клетку линейной двухнитевой ДНК фага Я она циркуляризуется за счет взаимодействия между липкими концами и превращается в ковалентно-непрерывную сверхспирализо-ванную форму. [12]
 |
Схема терминации синтеза ( 4 - цепи фага рХ174. [13] |
После попадания в клетку линейной двухнитевой ДНК фага А она циркуляризуется за счет взаимодействия между липкими концами и превращается в ковалентно-непрерывную сверхспирализо-ванную форму. [14]
Условия, необходимые для превращения линейной ДНК в кольцевую, заключаются в наличие у нее пары липких концов. Такие однонитиевые участки могут затем, аннелируясь, образовывать область дуплекса, давая кольцевую форму молекулы ДНК. Замкнутая кольцевая ДНК обнаружена в нативном состоянии во многих вирусах, бактериях и митохондриях, где эта форма ДНК является биологически активной [38], и может быть получена контролируемым химическим синтезом ( см. гл. [15]
Страницы: 1 2 3 4