Липкий конец
Cтраница 2
Необходимо расщепить две ДНК ре-стриктирующей эндонуклеазой, смешать и прогреть их, чтобы инактивировать эндо-нуклеазу, и разделить липкие концы в точках расщепления. Дать смеси остыть, с тем чтобы образовались нековалентные ре-комбинанты, после чего сшить их кова-лентно с помощью ДНК-лигазы. Кроме требуемого кольцевого дуплекса, в котором объединены исходные кольцевые ДНК, образуются побочные продукты: исходные кольцевые ДНК ( малая и большая) и две другие кольцевые ДНК, одна из которых является результатом рекомбинации двух больших ДНК, а другая-результатом рекомбинации двух малых ДНК. Кроме того, в смеси будут присутствовать линейные рекомбинанты, образованные двумя большими ДНК, двумя малыми ДНК, а также два типа рекомбинантов, состоящих из малой и большой ДНК, которые соединены в двух разных сочетаниях. [16]
Если такую ДНК выделить в чистом виде ( это умеют многие) и дать ей возможность свободно плавать в растворе, то липкие концы сомкнутся и ДНК превратится в кольцо. Поскольку эта молекула довольно длинная ( в ней около 40 000 пар нуклеотидов), то при замыкании в кольцо она с довольно высокой вероятностью завяжется в узел. [17]
Нагревание полинуклеотида LII при температуре несколько ниже температуры плавления и последующее медленное охлаждение приводит к образованию циклической структуры LIII за счет нековалентного взаимодействия комплементарных липких концов. Такая структура может быть обнаружена с помощью электронной микроскопии. Этот прием был использован для доказательства того, что концевые участки полинуклеотидных цепей ДНК фагов ТЗ и Т7 имеют одинаковую последовательность. Пользуясь сочетанием гибридизации и циклизации, можно различить вирусные ДНК, содержащие уникальную последовательность нуклео-тидов и представляющие набор циклически переставленных фрагментов274 ( см. стр. [18]
ДНК-лигазу фага Т4 можно применять и для сшивания ту-лых концов молекул ДНК, хотя этот процесс осуществляется труднее, чем в случае липких кондов. Липкие концы можно превратить в тупые путем отщепления одноцепочечных выступающих участков специфичными только к таким участкам нук-леазами. Альтернативный способ основан на использовании ДНК-полимеразы: она присоединяет к короткой цепи недостающие нуклеотиды, комплементарные 5 -одноцепочечной выступающей части молекулы. Лигаза применяется также для встраивания в плазмиды синтезированных фрагментов ДНК с целью получения на стыке ДНК-вставки и вектора нужной последовательности. Смысл этой процедуры состоит в том, что лповторное лигирование вектора становится невозможным. [19]

Такие молекулы смешивают с донорной ДНК, содержащей нужный ген и предварительно обработанной такой же рестриктазой. Поскольку липкие концы этих двух ДНК взаимно комплементарны, они спариваются с образованием гибридных молекул. Далее смесь обрабатывают ДНК-лигазой фага Т4 в присутствии АТР, в результате чего образуется множество разных комбинаций фрагментов, а также нежелательные продукты, в частности объединившиеся между собой фрагменты до-норной ДНК и исходные плазмидные ДНК. После репликации в трансформированной клетке одноцепочечные разрывы устраняются системой лигирования клетки-хозяина. [21]
Соединение соседних блоков ДНК водородными связями должно предшествовать ковалентному связыванию под действием лигазы. Эксперимент показал, что липкие концы должны содержать не менее 3 - 4 нуклеотидных звеньев. [22]
Соединение соседних блоков ДНК водородными связями должно предшествовать ковалентному связыванию под действием лигазы. Эксперимент показал, что липкие концы должны содержать не менее 3 - 4 нуклеотидных звеньев. [23]
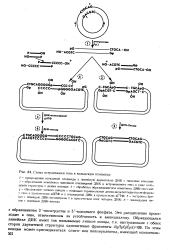 |
Схема встраивания гена в кольцевую плазмиду. [24] |
Это расщепление происходит в гене, ответственном за устойчивость к ампициллину. Образующаяся линейная ДНК имеет так называемые липкие концы, т.е. выступающие с обеих сторон двунитевой структуры однонитевые фрагменты d ( pTpCp CpA) - ОН. [25]
Для осуществления молекулярного клонирования недостаточно одних только ферментов рестрикции. Во-первых, водородные связи между теми четырьмя основаниями, которые образуют липкие концы, недостаточно прочны, чтобы удержать два объединившихся фрагмента ДНК. Этот фермент катализирует образование фосфо-диэфирных связей между концами полинуклео-тидных цепей, которые уже удерживаются вместе благодаря спариванию липких концов. [27]
Прн работе с рекомбинантнымн молекулами ДНК возникает необходимость замены одного типа липких концов на другой. Для этой цели используются адапторные линкеры - частично двухце-почечные нуклеотиды, содержащие липкие концы, соответствующие двум разным рестриктазам. Третий тип линкеров позволяет перейти от тупого конца ДНК к липкому, и их можно назвать дополняющими. [28]
Для получения линкеров синтезируют олигомеры, которые представляют собой палиндромные одноцепочечные нуклео-тидные последовательности, спаривающиеся ( гибридизующиеся) между собой. Разрезают новую молекулу нужной рестрицирую-щей эндонуклеазой и получают фрагменты с выступающими одноцепочечными концами ( липкими концами), с помощью которых встраивают ДНК-мишень в соответствующий вектор. Прежде чем проводить встраивание, рестрицированную смесь фракционируют для отделения ДНК с липкими концами от лишних линкерных молекул. ДНК-мишень не должна содержать сайтов рестрикции, присутствующих в линкерной последовательности, в противном случае она также будет расщепляться ферментом. [29]

Страницы: 1 2 3 4